
13.19 Introduction to endophytes
Many plants harbour fungi within their tissues that are at least harmless and may be beneficial. There are several definitions of endophytes but the emphasis is usually on the fact that although they live inside a plant, they do not cause any disease symptoms. The presence of endophytic fungi within symptomless plants is usually recognised indirectly through culturing methods: small pieces of tissue (leaf, for example) are surface sterilised, placed on agar plates, and the fungi inside the leaf grow out on to the agar.
Endophytic fungi can also be observed directly within leaves using histological methods with the light microscope. A particularly successful technique is to label one of the constituents of fungal cell walls with a fluorescent dye as this enables them to be observed in thin sections using a microscope equipped with fluorescence optics (Fig. 17).
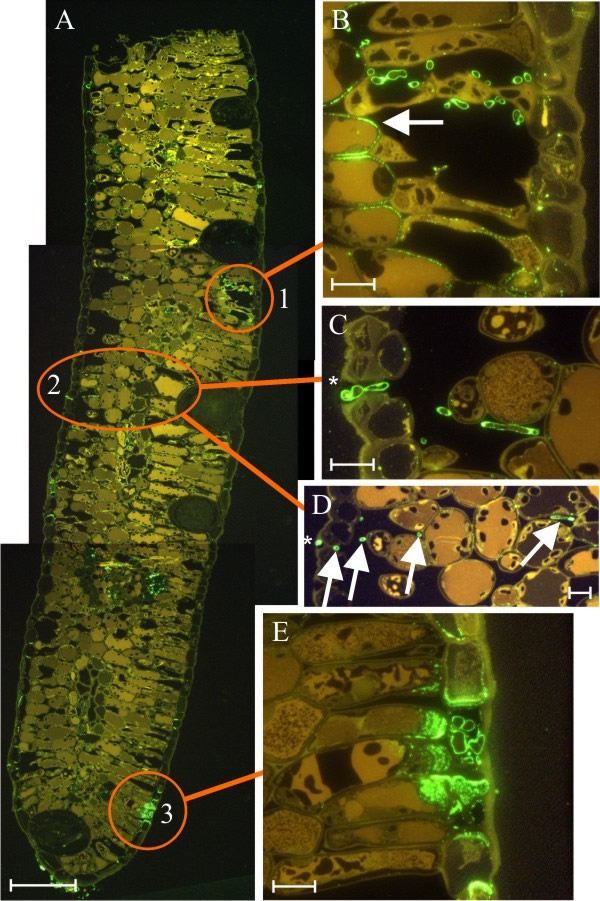 |
| Fig. 17. Section from a Kunzea ericoides leaf, 200 nm thick (panel A), with fungal cell walls fluorescently labelled to detect (1-3)-β-D-glucan (K. ericoides is the ‘white tea-tree’, a tree that occurs naturally in New Zealand). Walls of fungal hyphae label green, the plant cell walls autofluoresce to appear yellow or brown although plant cell walls with callose production also appear green. Three separate fungal infections occur in the photomontage that makes up the leaf section A, each of which is illustrated in detail in panels B to E. Panel B shows infection type 1, in which entry occurs through a stoma, fungal hyphae are restricted to the stomatal cavity, and the plant cells around the stomatal cavity produce callose (arrowed) in reaction to the fungus. Panels C and D show infection type 2, where entry again occurs through a stoma, but the plant cells surrounding the substomatal cavity show with no callose production, and fungal hyphae extend deeply into the leaf tissue (arrows in Panel D), asterisk indicates the same stoma in Panels C and D. Panel E shows the third type of infection, where the fungus directly penetrates the epidermal cell wall, and extensive callose production in surrounding plant cells restricts the fungus to a single epidermal cell. Scale bars: Panel A = 100 µm, Panels B–E = 10 µm. Modified from Johnston, Sutherland & Joshee (2006) using graphics files kindly supplied by Dr Peter R. Johnston, Landcare Research, New Zealand. Images reproduced with permission from Elsevier. |
These sections show that each plant host is usually colonised by several fungal species concurrently. They also show that the category ‘symptomless’ has different interpretations depending on the level of analysis. Microscopic examination may reveal that the plant is responding to an infection in an apparently healthy leaf that is symptomless to the naked eye (Johnston et al., 2006; Currie et al., 2014). Endophyte genome analysis has revealed the genes that are important for the endophytic life style, and further application of these approaches will explore the role of endophytes in host plant ecology and their biotechnological promise (Sanjana et al., 2016).
Fungal endophytes mainly belong to the Ascomycota and are a tremendously diverse range of fungi. They can be found in the above-ground tissues of all major lineages of land plants: liverworts, hornworts, mosses, lycophytes, equisetums, ferns, and seed plants. They range from the arctic to the tropics and occur in agricultural fields and the wilderness. There is another continuum in that they have a wide range of impacts on their hosts, ranging from the strictly symptomless cohabitant through to potential pathogens that can become pathogenic to their host plant in response to some change in circumstances. Within that continuum lies the possibility that the endophyte is symbiotic with its host but not all endophytes are mutualists (Arnold, 2007; Lacava & Azevedo, 2014). Over the last ten years metal hyperaccumulating flowering plants have attracted much research interest because of their potential use in clean-up of metal contaminated soils. Noccaea (or Thlaspi) caerulescens (Alpine Pennycress) has become the experimental model plant for studying Zn and Cd hyperaccumulation but recent work has demonstrated that endophytic fungi are responsible for enhancing the hyperaccumulation of Ni in this plant (Ważny et al., 2021).
Endophytes became a significant topic of study when those living entirely within grasses (in particular, tall fescue) were shown to be responsible for the toxicity of grasses to livestock. Fungal alkaloids produced by the endophyte are toxic to livestock that graze on the plant, but they improve the health of the plant by protecting against attack by grazing insects. So in this case the endophyte is conferring an advantage to its host. Unfortunately, it’s bad for farmed livestock so endophyte-infected tall fescue must be avoided in livestock feed, though it is actively encouraged for amenity sites, especially golf courses because the natural protection offered by the endophytic fungus means reduced use of chemical pesticides. Endophyte symbiosis in tall fescue can have a dramatic effect on many aspects of natural ecological systems; influencing plant-plant competition, diversity, productivity, succession, plant-herbivore interactions and energy flow through the food web (Rudgers & Clay, 2007; Omacini, 2014).
A functional relation to the host is not always obvious, many endophytes simply colonise inner spaces of plants like any other moist secluded habitat. The overall endophyte-plant interaction, though, depends on environment, and the age and genotype diversity of the host plants. Endophyte interactions of forest trees and other woody plants range from mutualism to pathogenicity (Saikkonen, 2007; Verma & Gange, 2014). Although mutualism is often assumed, the evidence is often circumstantial and the main reason for this is that endophytes are so common that endophyte-free control trees are simply lacking. Sieber (2007) summarised the situation like this:
‘…tree endophytes are mostly harmless colonisers of the internal [spaces] of healthy plant tissues. Some are potentially pathogenic but disease is only caused in combination with other, mostly unknown, inciting factors. Proof of mutualism of endophyte-host symbioses has been inconclusive in most cases, but plant communities would probably not survive many environmental stresses without these symbioses. All we know for certain is that endophytes are present in any healthy plant tissue!’
Endophytes have been evolving with their hosts for millions of years; and not just their hosts, they have been evolving with their host’s community and ecosystem, and that includes the pests and pathogens that prey on the host. Gall-forming insects are among the pests that infest plants and cause abnormal outgrowths (‘tumours’) of plant tissues formed in response to the stimulus caused by the insect laying eggs, or its larvae or nymph feeding on the plant. The insect becomes enclosed by the gall and feeds only on gall tissue during its development. These outgrowths are called galls because they contain large amounts of tannin which has a very bitter taste.
The insects involved include aphids, phylloxerans, psyllids, midges (gall gnats) and cynipid wasps (gall wasps) but the last of these, the gall wasps (order Hymenoptera, family Cynipidae) are the most important insects that induce plant galls, and about 80% of gall wasps produce galls specifically on oak trees (in fact, 60% of all known insect galls occur on oaks).
The endophytic fungi have been evolving with the oak and its gall wasps for a long time and a particularly interesting interaction has emerged for the oak gall wasp of Oregon white oak, because the fungal endophyte grows from the leaf into the gall and infects all the gall tissue. As a result, the insect in the gall dies but it is not directly killed by the fungus; rather, it dies because its food source, the gall tissue, is killed by the fungal infection. Approximately 12.5% of the galls on the tree die as a result of invasion by the fungal endophyte, so it’s a very considerable contribution to control of the pest. When the pest has been dealt with, the fungus returns to its life as a symptomless inhabitant of the leaf tissue (Lacava & Azevedo, 2014).
The status of most endophytes is still unclear but is evidently complex. For example, Fusarium solani strain K is a root endophyte of Solanum lycopersicum (tomato), which protects against root and foliar pathogens (Skiada et al., 2019). Fungal hyphae of strain K proliferate within the vascular bundle of the plant, and plant cell death is involved in fungal colonisation of the root. Establishment of Fusarium strain K within the plant tissues requires both fungal growth adaptations and plant cell-autonomous responses of the sort that occur during both symbiotic and pathogenic plant-fungal interactions. Fusarium solani strain K, endophytic in the legume Lotus japonicus, induced the expression in L. japonicus of several members of the common symbiotic signalling pathway (CSSP) that has been recorded as being triggered when symbiotic rhizobia and arbuscular mycorrhizas interact with legumes (Vasiliki et al., 2019; Skiada et al., 2020 ). The panel of induced expression included several plant receptors that perceive chitin molecules and have a role during recognition of Fusarium solani strain K by the plant. The results demonstrate that the CSSP is more widely employed in plant-microbe interactions than previously believed.
A variety of different factors can trigger the transformation of the fungus from innocent saprotroph, to mutualist, to pathogen. And they can be aggressive pathogens, often killing large parts of their host. But this usually follows physical damage or general stress on the host plant. There are even some observations that indicate structural integration of endosymbiotic fungi with the cells and tissues of the host, reminiscent of the lichen association (Barrow et al., 2007; Lucero et al., 2008).
Higher plants are ubiquitously colonised by fungal endophytes, and at a time when emerging dramatic climate changes are likely to increase stress on plant communities, endophytes might become the source of new plant diseases (Slippers & Wingfield, 2007), and could also be the source of valuable new secondary metabolites (Suryanarayanan et al., 2009; Higginbotham et al., 2013; Verma & Gange, 2014; Hodkinson et al., 2019; Abdel-Azeem et al., 2021). However, these ubiquitous hidden fungi may be important bodyguards of plants, protecting against grazing by insect herbivores (Gange et al., 2019).
Updated August, 2021