
13.18 Introduction to lichens
Lichens are mutualistic (symbiotic) associations between a fungus (the mycobiont) and a photosynthetic partner (the photobiont, also called the phycobiont as most are algal). It is usually an association between a fungus with a green alga, sometimes a cyanobacterium, as photobionts. The basis of the association is that the photobionts use sunlight to produce carbohydrate photosynthetically that is shared by both partners in the lichen. Some lichens contain both green algae and cyanobacteria as photobionts, in which case the cyanobacterium may focus on fixing atmospheric nitrogen for the joint metabolism of the tripartite association.
Using metatranscriptomics to examine the lichen symbiosis, Spribille et al. (2016) discovered that the classical binary view of lichens as a symbiosis between a single fungus, usually an ascomycete, and a photosynthesising partner, is too simple. They found that macrolichens are constituted of not two but three symbiotic partners: an ascomycete fungus, a photosynthetic alga, and, unexpectedly, a basidiomycete yeast. The yeast cells are embedded in the cortex of the lichen thallus and may contribute significantly to its morphology. The yeasts are ubiquitous and essential partners for most lichens and are not casual colonisers or parasites. And then there are the bacteria. In addition to nitrogen-fixing cyanobacteria, five bacterial orders are frequently found, providing functions to the overall symbiotic community of the lichen ranging from the provision of vitamins and cofactors to the degradation of phenolic compounds (Cernava et al., 2017). Hawksworth & Grube (2020) re-defined the lichen symbiosis as: ‘a lichen is a self-sustaining ecosystem formed by the interaction of an exhabitant fungus and an extracellular arrangement of one or more photosynthetic partners and an indeterminate number of other microscopic organisms’.
There are about twenty thousand species of lichens of all types in the living world (the current global checklist of lichens and allied fungi lists 18,882 species; Feuerer & Hawksworth, 2007). The lichen association is an intimate symbiosis which greatly extends the ecological range and scope of all partners. The lichen tissue is called a thallus, and in most cases, it is very different from either a fungal mycelium or an alga growing separately. The fungal hyphae surround the algal cells, often incorporating them into multicellular fungal tissues unique to lichen associations. They vary in size, shape and colour. Some are flat and firmly attached to the surfaces on which they grow, like those white or yellow-brown disks that are often seen scattered over walls and roofs. But others are scaly, leafy or bushy, or hang in strands from their supports. Several lichen thallus morphologies are recognised, and some are very similar to simple plants in appearance and growth pattern (Büdel & Scheidegger, 2008; Sanders, 2001):
- crustose (a flat crust, commonly seen on walls and other stonework);
- foliose (leafy);
- fruticose (branched, shrubby);
- filamentous (hair-like);
- leprose (powdery);
- squamulose (made up of small scales);
- gelatinous (lichens in which the cyanobacterium produce a polysaccharide that absorbs and retains water).
For illustrations see Brodo et al. (2001) [and visit the ‘Lichens of North America’ website at http://www.lichen.com/] Purvis (2007), and for more ‘floristic’ detail of biodiversity, Feuerer & Hawksworth (2007) and Smith et al. (2009). There are expectations that the increasing availability of molecular data will improve appreciation of lichen biodiversity and evolution (Lutzoni et al., 2018). However, generating genome-scale data from multi-species symbioses is challenging because metagenomic DNA from inseparable symbiotic organisms yields sequenced loci that may belong to any number of the organisms involved in these intimate associations. Nevertheless, appropriate approaches and methods are being devised (Aschenbrenner et al., 2016; Grewe et al. 2017; Meiser et al. 2017).
In microscope sections, a typical foliose lichen thallus (Fig. 15) reveals four zones of interlaced fungal filaments. The uppermost zone is formed by densely interwoven hyphae forming an outer protective tissue layer called the cortex, which might be a few hundred μm thick (Büdel & Scheidegger, 1996). In some Parmeliaceae the cortex secretes an epicortex, up to 1μm thick, which seems to be a sort of cuticular layer. The algal cells occur in a zone beneath the cortex embedded in a dense hyphal tissue. Each cell or group of cells of the photobiont is usually individually enveloped by hyphae. Under the algal layer is a third zone, called the medulla, of loosely interwoven fungal hyphae without algal cells. The lower surface of the thallus, beneath the medulla, is similar to the upper surface; it is called the lower cortex, and may consist of densely packed fungal hyphae. Hyphal branches emerging from the lower cortex, as root like structures known as rhizines, serve to attach the thallus to its substratum.
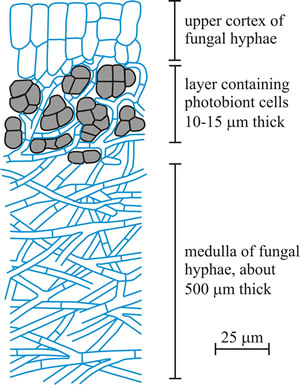 |
Fig. 15. A sketch of a slice of lichen, showing the general structure of a stratified (heteromerous) thallus. The majority of lichens develop internally stratified thalli; the most usual layers being an upper cortex, photobiont layer, medulla and, sometimes, a lower cortex. Lower surfaces, though, more usually lack a cortex, and are mycelial (described as ‘cottony’), often with fungal hyphae aggregated into strands so that the undersurface of the thallus appears to have a network of veins. The fungal tissues may be composed of elongated hyphae in a fairly loose mycelium (prosoplectenchymatous), or may be tightly packed so that individual hyphae are not distinguishable and the tissue seems to be made up of isodiametric cells, this is called pseudoparenchymatous tissue because it resembles the genuine parenchyma of vascular plants. The sketch shown illustrates a lichen like Peltigera, which is a foliose lichen with broad lobed thalli occurring in many parts of the world growing on soil, rocks, trees, and similar surfaces. All species of Peltigera associate with the nitrogen-fixing cyanobacterium Nostoc, which is probably the most widely distributed ‘blue-green alga’ in the world. Some also have a eukaryotic algal photobiont. Because of their dual abilities to fix nitrogen from the atmosphere and photosynthesise, such lichens are crucial primary colonisers and actually start the process of soil formation. For more details see Nash (2008a) and Sanders (2001). This figure redrawn and modified from an illustration in Hudson, 1986. |
The most ancient associations, in evolutionary terms, are between fungi and blue-green algae. Blue-green algae are better called cyanobacteria because they are bacteria rather than algae, but they do have chlorophyll and can photosynthesise. Cyanobacteria were the first organisms to release oxygen into the Earth’s atmosphere so they started, probably three billion years ago, the revolution from which emerged the atmosphere on which we rely today. The most ancient lichen-like fossils so far reported were found in South China. These fossils involve filamentous hyphae in close association with coccoidal cyanobacteria or algae. They are between 551 and 635 million years old, and indicate that fungi developed symbiotic partnerships with photosynthetic organisms before the evolution of vascular plants (Yuan et al., 2005).
Fossil lichens have also been described from the Rhynie Chert which is 400 million years old (Taylor et al., 2015; Krings et al., 2018). As the more advanced (eukaryotic) algae evolved the fungi (mostly relatives of present day Ascomycota) developed the partnerships that are now lichens. It has been suggested that lichenisation is ancestral and has been gained infrequently during the onward evolution of Ascomycota, and indeed, that the major Ascomycota lineages of non-lichen-forming species alive today might be derived from lichen-forming ancestors (Lutzoni et al., 2001, 2018).
Another entertaining evolutionary speculation you might like to read about is the suggestion that vascular plants originated from a genomic combination of fungal genome + algal genome providing the genetic architecture for a ‘higher plant body’ which is visualised as a mosaic of generalised alga-type cells (the photosynthetic cells of modern plants) interspersed with highly specialised fungus-type cells, which make up the structural and translocatory tissues (Atsatt, 1988). Stated like this, the notion may sound fanciful, but it has been regularly restated over the years (Jorgensen, 1993; Selosse & Strullu-Derrien, 2015; Aanen & Eggleton, 2017) and we recommend you read all these publications. Even more remarkable is the report that cells of the alga Nannochloropsis oceanica can, under certain conditions, become internalised within the hyphae of the fungus Mortierella elongata (Mortierellomycotina) (Du et al., 2019). In her discussion of some of the questions raised by this observation, Bonfante (2019) emphasises that prior to this observation in all known interactions between algae and fungi, notably all lichens, the algal cells remain outside the hyphae of the fungus. But in this study, which involved co-cultivation of alga and fungus, the algae first aggregated on the hyphal surface, and eventually algae entered the hyphae where they continued to photosynthesise, grow and divide. Nutrient exchange, including carbon and nitrogen transfer between fungal and algal cells was demonstrated by isotope tracer experiments. Both partners remained physiologically active over two months of co-cultivation. When the alga Nannochloropsis grows within the hyphae of Mortierella, those hyphae changed colour to green due to the photosynthetic activity of the algae within.
A comprehensive phylogenetic analysis of 259 transcriptomes and 10 green algal and basal land plant genomes concluded that the most recent common ancestor of present day land plants and green algae was preadapted for symbiotic associations, specifically by having the genes required for fungus-plant nutrient exchange. The study further suggests that subsequent rounds of gene duplication led to the acquisition of additional pathways, required for the perception and transduction of diffusible fungal signals for root colonisation that enabled formation of the arbuscular mycorrhizal symbiosis (Delaux et al., 2015). Could it be that the algal ancestor of land plants was ‘preadapted for symbiosis’ by being the photobiont in a lichen association?
For reproduction several lichens produce flakes comprising small groups of algal cells surrounded by fungal filaments (called soredia; Fig. 16) that form a powdery mass in structures called soralia on the upper surface of the lichen.
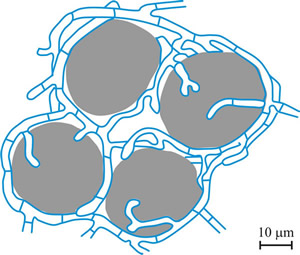 |
Fig.16. Soredium. Lichen reproductive structure composed of algal cells (shown in grey) enveloped by hyphae (shown in blue). Redrawn and modified from an illustration in Hudson, 1986. |
Other structures are fragile upright elongated outgrowths from the thallus (called isidia) that break off for mechanical dispersal. Many lichens break up into fragments when they dry, dispersing themselves by wind action, to resume growth when moisture returns; fruticose lichens in particular can easily fragment. In any of these cases the fragments include both fungus and alga and are easily dislodged to be blown about in the breeze like spores. They can begin a new colony if they land in a suitable place.
Lichens are the classic examples of symbiosis, in fact they prompted the term to be coined. Lichens dominate more of the land surface of Earth than do tropical rainforests, they involve one in five of all fungi, and can live in places that are inaccessible to other organisms. All this because of their ultrastructurally and physiologically unique partnership. The fungus protects the alga and supports it physically and physiologically by taking in water and using its externalised enzymes to extract nutrients from the soil, and even from the rocks themselves. These organisms directly or indirectly induce chemical weathering leading to the mobilisation of minerals like magnesium, manganese, iron, aluminium and silicon (Nash, 2008c). The algal or cyanobacterial cells in the partnership can photosynthesise; photolysing water and reducing atmospheric carbon dioxide to provide carbohydrates for use by both partners.
A parallel genomic analysis of the mycobiont Cladonia grayi and its green algal photobiont Asterochloris glomerata in coculture showed that the fungus upregulated small secreted proteins, membrane transport proteins, signal transduction components, extracellular hydrolases, a ribitol transporter and an ammonium transporter, while the alga activated DNA metabolism, signal transduction, and expression of flagellar components. Expanded fungal protein families included heterokaryon incompatibility proteins, polyketide synthases, and a unique set of G-protein subunits. Expanded algal protein families included carbohydrate metabolism enzymes and a specific subclass of cytoplasmic carbonic anhydrases. The symbiosis-related genes affected stress resistance, signalling, genome reprogramming, and nutritional and structural interactions. The diversity of the genes involved is consistent with the idea that ascolichens could have had a few independent evolutionary origins (Armaleo et al., 2019).
Questions have been raised about how truly mutually beneficial the relationship might be, however. Most of the body of the lichen is fungus; the algal partner making up only five to ten percent of the total biomass. Indeed, microscopic examination shows that fungal cells might even penetrate the algal cells in a way similar to pathogenic fungi; although for the most part the readiness with which nutrients leak out of algal cells makes penetration by fungal haustoria completely unnecessary. Certainly, the photosynthetic partner can exist independently of the fungal partner in nature, but not vice versa.
If cultivated in the laboratory without the photobiont, a lichen fungus grows as an undifferentiated mass of hyphae. Only when combined with its photobiont does the hyphal morphology characteristic of the lichen thallus arise. Cells of the photobiont are routinely destroyed in the course of nutrient exchange; stability of the association depending on photobiont cells reproducing more rapidly than they are destroyed. Because of these observations it has been suggested that the fungus in the lichen is really parasitising the alga and using the products of algal photosynthesis for fungal nutrition (Palmqvist et al., 2008). However, other observations support the notion that ‘mutualism’ best describes the association, particularly the numerous metabolites that the two partners share, and the relationship has been likened to a farming operation in which the fungus is ‘farming’ its photobiont. The statement ‘Lichens are a case of fungi that have discovered agriculture...’ has been attributed to the lichenologist Trevor Goward (Department of Botany Herbarium, University of British Columbia) (Sanders, 2001).
Lichens have a remarkable ability to thrive where no other organisms can exist. They are often the first to settle in places lacking soil, constituting the sole vegetation in some extreme environments. A characteristic feature is that they can tolerate severe desiccation. In dry conditions lichens go into metabolic stasis (known as cryptobiosis) where dehydration of the cells of the lichen halts most biochemical activity. In this state lichens can survive extremes of temperature, irradiation and drought in the harshest environments (Honegger, 1998).
Lichens tolerate temperature extremes from the heat of deserts to the cold of Arctic and Antarctic wastes. Some grow on high mountain rocks, extracting moisture from mists and fog. They are also extremely common as epiphytes growing on the surfaces of plants, particularly on the trunks, branches and even twigs of trees. In these circumstances the epiphytic lichens are not parasites; they have no physiological impact on the plant; they are simply using its surface as a habitat. Understandably, lichen growth can be rather slow; maybe two inches per thousand years in the Arctic but somewhat faster in less extreme environments.
About 13,500 species of fungi are involved in lichens, representing about 20% of all known fungi. They are mainly Ascomycota with a few (only about 15) Basidiomycota, which are called basidiolichens to differentiate them from the much more common ascolichens. Even though lichens are characteristically found in environments subject to temperature, desiccation and low nutrient stresses, fungal sexual structures are often formed in abundance, a feature which obviously contributes to their diversity and ecological success (Seymour et al., 2005). This may be a common form of reproduction in basidiolichens, which form fruit bodies similar to their nonlichenised fungal relatives. In the ascolichens, spores are produced in apothecia, perithecia and pycnidia. After dispersal, these fungal spores must find a compatible algal partner before a functional lichen can be formed.
The taxonomic naming of lichens is based on the fungal component. As lichens are resistant to environmental extremes they are pioneer colonisers of terrestrial habitats, such as rock faces, tree bark, roofing tiles, etc., so they are very common in both urban and rural environments (Purvis et al., 1992), where lichen species diversity can be used as a measure of atmospheric pollution (Nash, 1996). Lichen dependence on the atmosphere and rainfall makes them highly sensitive to atmospheric pollution. Acidic rain and sulfur dioxide kill many lichens, so cities in industrialised countries may have few lichens because of the poor air quality. So if you do have lichens growing on your stonework it’s probably good for your lungs!
The nutritional value of lichens is similar to cereal seeds, though they do not make major contributions to human food. In the harsh areas where lichens grow, native peoples use them as food supplements. One lichen, which occurs in the deserts of the Middle East, may have been the manna that fell from heaven to rescue the Children of Israel from starvation in the Old Testament story:
“...and in the morning the dew lay round about the host. And when the dew that lay was gone up, behold upon the face of the wilderness there lay a small round thing, as small as the hoar frost on the ground. And when the children of Israel saw it they said one to another, It is manna: for they wist not what it was. And Moses said unto them, This is the bread which the Lord hath given you to eat...” [Exodus, 16, v. 13-15].
Lichens are much more important as animal food, especially in the Arctic, where lichens can form as much as 95% of the diet of reindeer. In less harsh conditions, most mammals of these wildernesses will supplement their normal winter diets with lichens. Many Lepidopteran larvae feed exclusively on lichens. In hotter climates, sheep in Libya graze a lichen that grows on the desert rocks. Grinding their teeth away as they chew it off. The extreme life style of lichens leads them to produce several exotic chemicals, some of which are very useful for lichen identification. These compounds may be useful to humans. They include antibiotics, essential oils for perfumery and dyes for textiles, and there are probably many others awaiting discovery and exploitation (Oksanen, 2006; Elix & Stocker-Wörgötter, 2008; Cernava et al., 2015).
Updated July, 2020