
13.11 Ericoid endomycorrhizas
Most plants belonging to the order Ericales are able to associate symbiotically with soil fungi to form the distinctive ericoid mycorrhiza. This association was initially described in members of the family Ericaceae, which is more abundant in the northern hemisphere and is probably best known in the UK through heather moorland (‘heathland’) genera characterised by Erica (heather), Calluna (ling) and Vaccinium (bilberry), though the group also includes Rhododendron. These are plants that endure moorlands and similar challenging environments, as heathland habitats are typically found at high altitudes and colder latitudes, and have nutrient poor, acidic soils. A morphologically similar mycorrhizal association occurs in the family Epacridaceae, which is widely distributed in the southern hemisphere, particularly southern Australia; the family takes its name from the genus Epacris.
 |
|---|
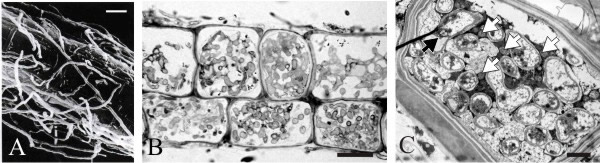
| Fig. 9. Cell biology of ericoid mycorrhizas. A. Scanning electronmicrograph of Rhododendron root covered with a loose weft of fungal hyphae; modified from Smith & Read, 1997. B, section of the ericoid mycorrhiza of Gaultheria poeppiggi (Ericaceae); the cortical cells are well colonised by fungal hyphae, scale bar = 10 μm. C, cortical cell of an Erica cinerea hair root colonised by hyphae (they make up most of the volume of the cell): black arrowhead = a dolipore septum of an intracellular hypha; white arrowheads indicate mitochondria of the plant cell, scale bar = 2 μm. B and C modified from Selosse et al. (2007) using graphics files kindly provided by Prof. Marc-André Selosse, Université Montpellier II, France and Dr Sabrina Setaro, Wake Forest University, NC, USA. Reproduced with permission of John Wiley and Sons. |
The fungi involved in ericoid associations are members of the Ascomycota. Ericoid fungal hyphae form a loose network over the hair root surface (Fig. 9A). The hyphae also penetrate the epidermal cells, often at several points in each cell (Fig. 9B, C), and those cells become filled with coils of hyphae. As in all endosymbioses, the intracellular fungal symbiont is separated from the plant cytoplasm by a plant-derived membrane, which invaginates to follow fungal growth and coil formation. Up to 80% of the hair root volume can be comprised of fungal tissue. It is through these coils that nutrient exchange occurs. This colonisation by the fungus is limited to mature and fully expanded epidermal cells. In the apical region of the hair root, just behind the growing meristem, the immature epidermis remains uncolonised.
Ericoid mycorrhizal fungi are facultative symbionts; that is, they can exist as free-living mycelia in the soil, and they can also be cultured in artificial media. When grown on nutrient agar the fungi produce dark-coloured, slow growing and sterile mycelia. Absence of spores and reproductive structures has complicated the identification and classification of ericoid fungi and generated increasing use of molecular techniques for examining DNA and RNA profiles. These show that there is considerable genetic diversity between isolates that are superficially similar in appearance. Comparative genomics and transcriptomics show that the fungal genes most highly upregulated in the ericoid mycorrhizal symbiosis are those coding for fungal and plant cell wall-degrading enzymes, lipases, proteases, and transporters. This gene transcription pattern suggests a versatile dual saprotrophic and biotrophic lifestyle quite like fungal endophytes (see Section 13.19; Martino et al., 2018).
Ericaceous shrubs can become dominant in many heathland communities, especially in environmental conditions where plant litter is only slowly decomposed, resulting in acidic soils rich in hard-to-digest organic matter but low in available mineral nutrients such as nitrogen and phosphorus. These mycorrhizas improve nitrogen and phosphorus uptake by the plant, enabling the host plants to access otherwise unavailable organic nutrients. The network of hyphae covering the root and the mycelium in the soil digest polypeptides saprotrophically and absorbed nitrogen is exchanged with the plant host. In extremely harsh conditions (for example winter in high altitude and northern latitudes) the mycorrhiza may even support the host plant with carbon nutrients (by metabolising polysaccharides and proteins for their carbon content). Normally, though, the fungus takes photosynthetically-produced carbohydrates from the plant host.
These physiological attributes of the mycorrhizal fungus enable ericaceous plants to act as pioneer species, colonising unpromising habitats ranging from arid sandy soils in the southern hemisphere to moist mor humus (a raw humus state of unincorporated organic material, poorly mineralised and with an acid pH; only slightly more degraded than peat) of the northern hemisphere; and the absence of ectomycorrhizal fungi seems to prevent colonisation of heathland by tree species like pines and birch (Collier & Bidartondo, 2009). Also thanks to the mycorrhizal fungi, ericaceous plants are able to grow in highly polluted environments, where the soil contains otherwise toxic levels of metal ions. Taken together, the mycorrhizal symbiosis enables ericaceous plants to survive under nutrient stressed conditions and improves the ability of the host to compete successfully with other plants even on polluted sites contaminated by heavy metals (Mitchell & Gibson, 2006; Perotto, Girlanda & Martino, 2002).
Updated July, 2019