










5. Quantitative analysis of cell size and distribution
Most mycologists must be well aware of the description of tissue construction in mushrooms and toadstools which is called hyphal analysis. This is a procedure whereby the range and type of differentiated cell types are catalogued and used as taxonomic criteria. Hyphal analysis was introduced by Corner (1932a; 1932b), who coined the terms monomitic, dimitic and trimitic to describe tissues consisting of one, two or three kinds of hyphae. Later, the words sarcodimitic and sarcotrimitic were used to describe fruit bodies having two or three types of hyphae of which one is inflated and has thickened walls (Corner, 1966; Redhead, 1987).
Hyphal analysis in this sense is an almost entirely descriptive study, aimed at establishing structural features as taxonomic criteria. Many of the cell types which hyphal analysis usefully identifies are named for their morphological features alone (the essence of taxonomic description), yet the names carry functional overtones (words like 'generative' and 'skeletal' are used) though this is entirely a matter of presuming a function without proof or even evidence beyond the morphology. Walls which are seen to be unusually thick by light microscope observation are almost always assumed to be mechanically strengthening (= skeletal or ligative hyphal characters). Yet fungal wall structure is modified both chemically and physically to serve as a transient nutrient store, so wall appearance is not a reliable guide to wall function. Nevertheless, the taxonomic importance of hyphal analysis is immense (Pegler, 1996). It can even be applied in vitro to identification of fungi (especially wood rotting species) in culture (Nobles, 1958, 1965, 1971; van der Westhuizen, 1958, 1963, 1971; Stalpers, 1978; Rajchenberg, 1983; Lombard, 1990; Nakasone, 1990). My interest, however, is in the contribution it makes to our appreciation of two features. First, the analysis demonstrates the range of functional differentiation of which the hypha is capable. Second, the varied appearance of the hyphal types and the intergradations between them illustrates the adaptability, and versatility, the fungi express in constructing their fruit bodies.
The functional and morphogenetic purposes of the hyphal differentiation which hyphal analysis describes have only rarely been considered. Listing the presence or absence of a cell type is considered adequate. Consideration of its function, how it arises, how its position may be regulated, and its prevalence are seldom considered. The first quantitative hyphal analysis was done by Hammad, Ji, Watling & Moore (1993a) and Hammad, Watling & Moore (1993b) who showed that enumerating cell types at different stages of development (in the fruit bodies of Coprinopsis cinerea) is a powerful way of revealing how the macroscopic aspects of fruit body structure emerge during morphogenesis as a result of changes in hyphal type and distribution.
Details of the procedure, which uses computer-aided image analysis of microscope images, appear in Hammad et al. (1993a & 1993b). They discovered that the stem of Coprinopsis cinerea comprises two cell populations and that cell inflation is accentuated in cells occupying a specific zone of the stem. Differential expansion of cells in this zone readily explains how the stem changes from a solid cylinder to a hollow tube during its development.
Narrow and inflated hyphae in the fruit body stem of Coprinopsis cinerea. Although Coprinopsis is one of the most frequently studied genera there is surprisingly little information concerning the structure of the stem of its fruit body other than that the stem is composed of greatly inflated and elongated cells. Most observers would consider the main body of the stem to be basically uninteresting, being made up exclusively of inflated cells. Hammad et al. (1993b) measured cross-sectional areas of hyphal profiles in 5 μm thick transverse sections cut from stems of fruit bodies of a range of developmental ages. For each section of each piece of stem, the area of every cell within two randomly chosen radial transects 12 μm wide was measured. On the basis of frequency distributions two distinct populations of hyphae were identified and categorised as narrow hyphae, with cross-sectional area <20 μm2, and inflated hyphae, with cross-sectional area ≥20 μm2. Narrow hyphae constituted 23% to 54% of the cells in transverse sections of the stem but only contributed 1% to 4% of the overall cross-sectional area.
All the available evidence indicates that narrow hyphae in stems of Coprinopsis cinerea have diverse functions. For one thing they react differently to a range of standard histological stains. Also, they tend to be particularly concentrated at the exterior of the stem where they may serve as an insulating layer, and as a lining to the lumen where they may represent the remnants of the initially central core of dikaryotic hyphae.
What makes some hyphae become inflated and multinucleate while others remain morphologically similar to the vegetative mycelial hyphae is not known although the even (i.e. non-random) distribution of the former (see Hammad et al., 1993b) implies some form of organisational control. This differentiation occurs at an extremely early stage as both narrow and inflated hyphae can be seen in primordia 3 mm tall. During stem elongation, however, the numerical proportion of narrow hyphae decreased. About 25% of the narrow hyphae were recruited to the inflated category as the fruit body developed.
The cell measurements of Hammad et al. (1993a) in Coprinopsis cinerea revealed little increase in cell length between a 3 mm fruit body and an 8 mm tall fruit body (both of which were at pre-meiotic developmental stages). Presumably any stem elongation occurring at these stages is due primarily to cell proliferation rather than cell elongation. By contrast, there was a large increase in cell length between the stems of the 8 mm fruit body (pre-meiotic) and that of a 25 mm fruit body undergoing meiosis. Initially the cells in the basal and middle regions of the stem lengthened. Cells in the extreme basal and apical regions were always shorter than those in other regions of elongated stems. For example, cells near the cap/stem junction at the extreme apex of an 83 mm fruit body (fully elongated) had a typical length of 150 μm compared with an average for the whole of the apical section examined (about 10 mm long) of 313 μm. The most elongated cells were found in the upper mid-region of the stem. Ratios of length to width were about 2 in pre-meiotic stems (3 mm and 8 mm fruit bodies), but increased after meiosis, particularly in the upper middle regions, to 10, 20 and approximately 35 in 48 mm, 55 mm and 83 mm tall fruit bodies respectively. Overall, therefore, developmental stem extension of C. cinerea involves increase in length and cross-sectional area of inflated hyphae and recruitment of narrow hyphae into the inflated population.
Developmental changes in the distribution of hyphal sizes within the stem. Comparing cell size with absolute position within transects of fruit bodies of different size revealed a progressive change in the distribution of inflated hyphae. In 6 mm and 27 mm tall fruit bodies the inflated hyphae increased in cross-sectional area up to halfway across the cortex but then their size declined towards the centre. In the 45 mm tall fruit body the cross-sectional area of inflated hyphae increased gradually from the exterior to the centre, and this pattern was even more pronounced in the 70 mm tall fruit body, where the peak cell area was adjacent to the lumen rather than in the mid-cortex (Fig. 27).
|
| Fig. 27. Interpretation of the geometrical consequences of the cell size changes during development of the stem of Coprinopsis cinerea. The graphs at the top of the figure show the lines of best fit for data relating hyphal cross sectional area to position across radii of 27 and 70 mm stems, together with scale drawings of the fruit bodies. The diagrams at the bottom of the figure are transverse sections of the stems, drawn to scale. The diagrammatic transverse section of the stem of a 27 mm tall primordium (bottom left) is composed of solid tissue which is divided into four zones corresponding to the zones in the radius in the graph above. The central (zone 4) and outermost (zone 1) zones are comprised of rather smaller cells than the two cortical zones. During further vertical growth the most dramatic cell inflation occurs in the cells of zone 3 which are here shown shaded. Growth from 27 mm to 70 mm in height is accompanied by a 3.6-fold increase in cell area in zone 3 and a 1.6-fold increase in zone 2. The cells in the other zones have to be rearranged to accommodate the inflation of zone 3 and a major consequence is that a lumen appears in the centre of the stem. Changes in area are shown to scale in these diagrams, though only a total of 327 cell profiles are illustrated. As this is only a tiny proportion of the cells involved in vivo, the diagrams inevitably distort the apparent relationship between cell size and stem size. Narrow hyphae have been ignored in these diagrams, though they are distributed throughout the tissue and their conversion to inflated hyphae contributes to stem expansion. From Moore, 1996a. |
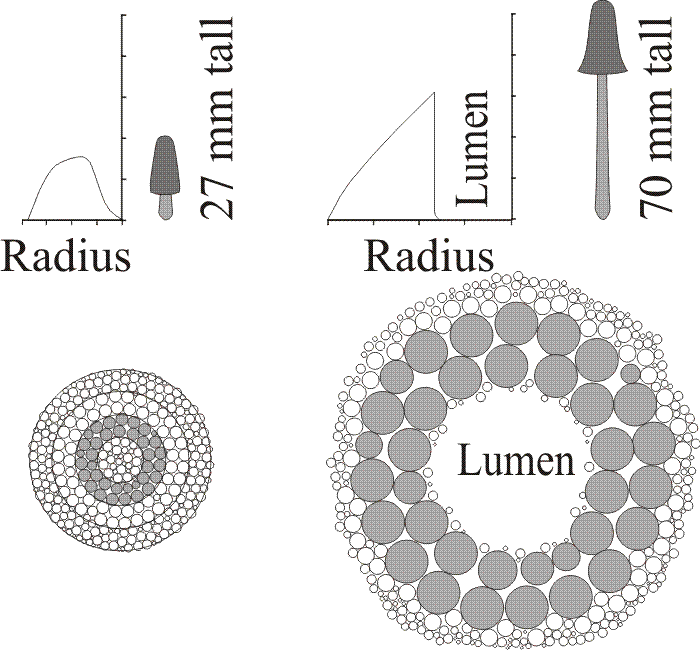
These data obtained by Hammad et al. (1993b) show that expansion of the stem is mainly due to increase in cross-sectional area of inflated hyphae in the region internal to the mid-cortex. Inflated hyphae around the periphery of the stem do not enlarge much. The mechanical consequences of this pattern of cell inflation are obvious. Increase in cross-sectional area of inflated hyphae in such a deep-seated zone will: (a) result in the central axis being torn apart, leaving its constituent cells as a remnant around the inner wall of the lumen so created; and (b) reorganise and stretch the tissues in the outer zones of the stem (Fig. 27). The outcome of these processes is a hollow cylinder with an outer 'skin' stretched over inner compressed tissue. In engineering terms, an ideal structure for a columnar support.
Consequently, formation of the mature stem in its optimum mechanical construction is entirely a result of a specific pattern of cell inflation within the stem as it develops. This particular pattern of inflation must be organised by signalling molecules which determine differential cell inflation across the stem radius (Moore, 1993).
Such observations concentrate attention on the integration of control of cell expansion throughout the fruit body. Expansion of the different cell types in the cap as well as inflation of cells of the stem began immediately post-meiotically (Hammad et al., 1993a). Such co-ordination may be achieved by some sort of signalling system that 'reports' the end of meiosis to spatially distant parts of the fruit body. The route such a signal might take is not clear, but primary gills are attached to the stem, with their tramal regions in full hyphal contact with stem tissues, so the connection between tissues undergoing meiosis and the upper (most reactive) regions of the stem may be fairly direct.
The overall implications of these studies are that positional information in otherwise homogeneous cell populations is imparted by some sort of signalling molecule(s) and that such signalling may occur over short (<μm) and long (>mm) distances (Moore, 1993). [TOP]
Updated December 7, 2016