
7.6 Biology of incompatibility systems
It is the compatibility reactions (including, for the moment, both vegetative compatibility discussed here and the mating type systems dealt with in Chapter 8) that define in real life what constitutes the fungal individual. In yeasts each cell is clearly an individual but a mycelial individual is not so obvious. Spores are individuals and colonies developed from single spores must also be individuals. But are ten spores from the same colony ten different individuals, or just ten bits separated from one individual? And then there are heterokaryons; mycelia that contain more than one nuclear type. Is a heterokaryon an individual, rather than a chimera or mosaic? These are important questions because in genetical terms a population consists of individuals that are able to interbreed. Individuals are important in evolution because selection operates on individuals. So to understand fungal populations we have to know where the individual begins and ends.
Populations are important because the fundamental unit of biological classification, the species, is conventionally defined in terms of mating success and production of viable offspring. This is the biological species (or genospecies) [CLICK HERE to view our Resources Box about the Species Concept]. Compatibility systems maintain the individuality of a mycelium and enable it to recognise unrelated mycelia of the same species with which it competes for territory and resources. In other words they provide an individual mycelium with a way of establishing whether hyphae it encounters belong to itself or not; fungi have many ways of expressing their individuality because individuality is of prime importance to fungi by reducing the spread within a species of harmful cytoplasmic mutations and/or viral or plasmid infections, and possibly being the first step towards speciation.
Fungal individuals are most clearly seen in dead tree stumps that are colonised by several mycelia of a wood decay species. Each mycelium explores the timber and attempts to capture a volume of the woody substrate for its own use. The mycelia interact where they meet and vegetative compatibility systems come into operation. If the strains are incompatible, demarcation zones of dead hyphae are formed between adjacent strains resulting from the death of fusion cells, killed by vegetative incompatibility. The living hyphae near these zones often form thickened and pigmented walls, which emphasise that the two mycelia are distinct. If the timber is sawn into segments these pigmented zones appear as pigmented lines (‘demarcation’ or ‘zone’ lines) on the cut surface. If the distinct zones extend to a surface at which they can form fruit bodies, a collection of fruit bodies expressing the different genotypes represented in that local population will appear (Fig. 8).
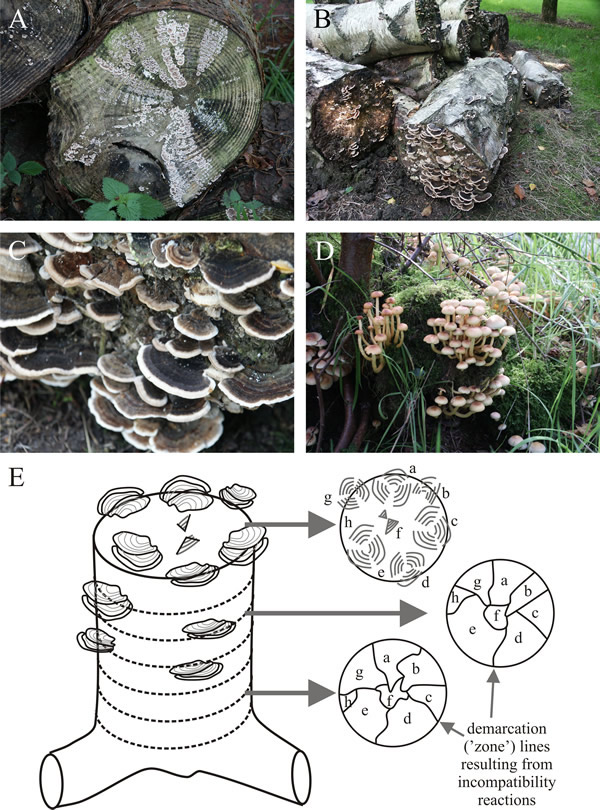 |
| Fig. 8. Felled logs colonised by mycelia of Trametes versicolor (Basidiomycota; commonly called Turkey Tail in the United States) (A, B, C) and Hypholoma fasciculare, D, commonly known as the Sulphur Tuft. Early in the season the mycelia reach the end of the log and the differentiating fruit bodies outline the separate decay columns in the timber (A), which are formed by mycelia belonging to different compatibility groups. Fruit bodies are formed on these surfaces later in the season (B, C and D). D, (Hypholoma fasciculare) shows that the formation of fruit bodies from distinct decay columns is not limited to Trametes, though in this specimen the log is completely covered in moss. The diagram shown in E is a tree stump colonised by eight incompatible mycelia of T. versicolor in which the decay columns within the timber have been mapped in successive sections (of about 1 cm thickness). The decay columns remain distinct because operation of the vegetative compatibility system produces a demarcation zone of dead and pigmented hyphae between each pair of adjacent strains. In sections of the timber these appear as demarcation (or zone) lines showing the boundary of each mycelium (a, b, c, d, e, f, g & h) (and see Fig. 13.4). Where a mycelium reaches the surface and fruits, it produces fruit bodies with the genotypes of that mycelium. Photographs by David Moore of specimens in the Lovell Tree Collection Arboretum at Jodrell Bank Discovery Centre, Cheshire (http://www.jodrellbank.net/); E based on Fig. 3 in Rayner & Todd, 1982. |
Updated July, 2019