
7.3 Breakdown of a heterokaryon
Unless there is a continual selection pressure favouring their continued existence, there is a tendency for heterokaryons to break down by forming sectors of homokaryotic growth. The nuclei of hyphae at the margins of heterokaryons are predominantly from one parent or the other, and if conditions change to favour one nuclear type, hyphae containing that nucleus will be selected (Fig 3).
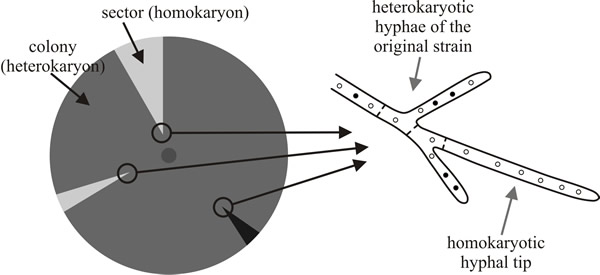 |
| Fig. 3. Sectoring within a heterokaryotic colony. ‘Sectored’ refers to the pattern of hyphae in a fungal colony on a Petri dish; sectors of homokaryotic hyphae are formed like the sectors of a pie-diagram. If the homokaryon and heterokaryon differ morphologically (for example by the colour of the spores they make), then these sectors are visible to the naked eye (diagrammed at left). The sectors arise when a hyphal tip receives nuclei of only one type and can continue to grow as a homokaryon (diagrammed at right). This may happen at any time in the growth of the colony; the closer it happens to the inoculum, the larger and more extensive is the final sector. Sectoring may be purely a matter of chance, depending only on the random distribution of nuclei between newly formed hyphal tips; but may be made more frequent by chemicals that disturb nuclear distribution or by selection pressure that favours the homokaryon. When there is no positive selection pressure to maintain a heterokaryon it is more likely to breakdown into its component homokaryons. For example, if the heterokaryon shown in Fig. 7.2 is grown on a ‘complete’ medium containing lysine and adenine, the absence of selection pressure favouring heterokaryon maintenance results in its breakdown into the component auxotrophic homokaryons. |
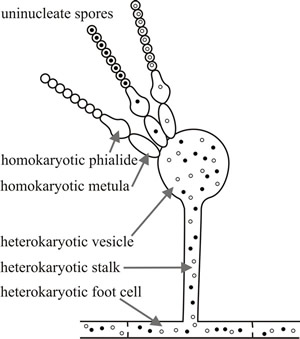 |
Fig. 4. Heterokaryon breakdown occurs as a result of asexual sporulation if uninucleate spores are formed, the uninucleate spores then germinate to form homokaryons. In Aspergillus nidulans, for example, the conidia are produced at the apex of the phialides (spore mother cells) on the sporangiophore vesicle. Each phialide contains only one nucleus which has migrated from the vesicle. This undergoes successive mitotic divisions; after each mitosis one daughter nucleus migrates into the developing spore and the other remains in the phialide for the next round of mitosis. Thus, all of the nuclei in the chain of uninucleate conidia formed from a single phialide are genetically identical. See Fig. 9.9 for more detail about the structure of the conidiophore of Aspergillus nidulans. |
Heterokaryons also breakdown as a result of asexual sporulation if uninucleate spores are formed, as in Aspergillus nidulans (Fig. 4); the uninucleate spores then germinate to form homokaryons.
In contrast, many fungi produce multinucleate spores, and if the mycelium producing these is a heterokaryon, the spores will also be heterokaryotic and the state will be propagated through asexual sporulation (an example of this is Botrytis cinerea, a pathogen of many plants and cause of the ‘noble rot’ of wine grapes, Fig. 5).
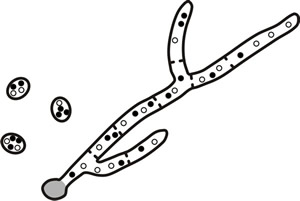 |
Fig. 5. In some fungi that produce multinucleate spores, heterokaryons can be propagated through germination of a heterokaryotic asexual spore (for example, Botrytis cinerea). |
Updated July, 2019