
15.4 Agriculture in beetles
The final example of an intimate interdependent association between an insect and a fungus concerns wood-boring beetles in the weevil subfamilies Scolytinae and Platypodinae, which are unusual in that the adults burrow into the trunks of living trees which have been under some sort of stress (drought, air pollution, etc.) for feeding and egg-laying; they may also be found in trees that have been recently cut or blown down. Rather than create fungal gardens underground like ants and termites, these female beetles dig tunnels and galleries into wood (Fig. 4), laying eggs on the tunnel wall and inoculating the wood with fungal material carried from a previous nest. They bring the fungi to their new host tree in one of several glandular small pits on the surface of their body, called mycangia or mycetangia.
Ambrosia beetles are completely dependent on their fungal symbionts and have deep and complicated pouch-like mycangia. These specialised structures on the body of the animal are adapted for the transport of symbiotic fungal spores and mycelia in pure, often yeast-like, cultures, secreting substances to support fungal spores and perhaps to nourish mycelium during transport. Once introduced, the fungi grow in the wood as mycelia and these help the beetle to evade tree defences as well as serving directly as food (Farrell et al., 2001). They are found in many xylophagous (‘wood-eating’) insects, although, despite this name, the beetles derive most of their nutrition from digestion of the fungi growing on the wood (Filipiak, 2018). In some cases, as in ambrosia beetles, the fungi are the sole food, and the excavations in the wood are only to create suitable microenvironments for the fungus to grow. In other cases (for example, the pine beetle, Dendroctonus frontalis) the fungus is carried by mites that ride on the beetles.
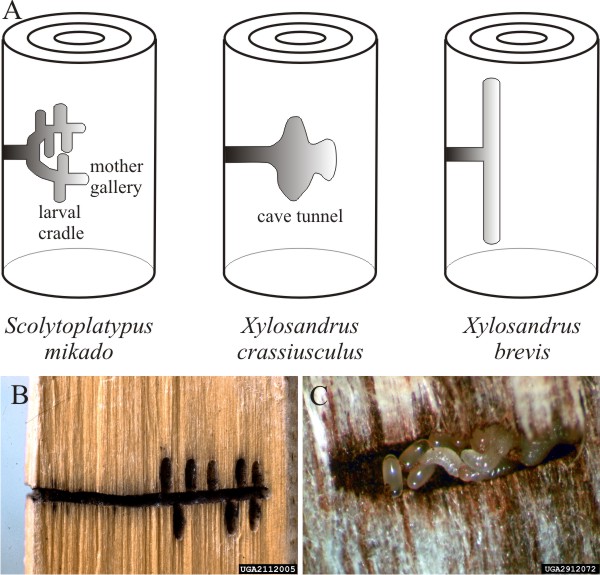 |
| Fig. 4. Ambrosia beetles are pests which damage timber. A, across the top, shows a set of diagrams of gallery structures of typical ambrosia beetles, with the names of beetles forming that type of gallery indicated below the diagrams. B, galleries of the striped ambrosia beetle (Trypodendron lineatum) in timber of Norway spruce (Picea abies)(photograph is image number 2112005 by Petr Kapitola, State Phytosanitary Administration, Department of Pest Risk Analyses, Czechia (http://www.forestryimages.org/browse/autimages.cfm?aut=11449). C, eggs and larvae of the granulate ambrosia beetle (Xylosandrus crassiusculus) (image number 2912072 by Will Hudson, Department of Entomology, University of Georgia, USA (http://www.forestryimages.org/browse/autimages.cfm?aut=153). Photographs B and C from Forestry Images (https://www.forestryimages.org/), a joint project of The Bugwood Network and USDA Forest Service; reproduced with permission. |
By the time the eggs hatch, the fungus will have grown over the tunnel walls, using its enzymes to digest constituents of the wood. This fungus ‘lawn’ (called, rather fancifully, ‘ambrosia’) provides the developing young larvae with a readily digested food. Ambrosia beetle larvae do little tunnelling; instead they feed together in chambers on asexually produced fungal conidia induced in cultures kept pure by the parents. Eventually, the larvae pupate and subsequently emerge as adults with a supply of fungus in their mycangia. Because they are ‘gratuitous’ food for the larvae, the fungi have become known as ambrosia fungi and the insects as ambrosia beetles.
Ambrosia fungi are derived from plant pathogens in the group known as ophiostomatoid fungi (that is, they are related to, or belong to, the genus Ophiostoma (order Ophiostomatales, subclass Sordariomycetidae, and phylum Ascomycota); the fungal cultivars are now classified to the genera Ambrosiella and Raffaelea. Other beetles in this group are known as bark beetles and are associated with free-living, pathogenic ophiostomatoid fungi that aid beetle attack of phloem of trees with resinous defences. In these cases the beetle is acting as a vector for the disease fungus; we have already described one such association in our discussion of Dutch Elm Disease (see the section entitled Ophiostoma (Ceratocystis) novo-ulmi (Dutch Elm disease or DED) (Ascomycota) in Chapter 14; CLICK HERE to view the page).
Ambrosia beetles total about 3400 species, and many have been found in 30 million-year-old amber, suggesting that the origins of this association occurred in the Tertiary, up to 60 million years ago, roughly contemporaneous with the origin of the attine ant symbiosis. However, in contrast to the single instance of origin of the attine ant symbiosis, there are at least seven independent instances of the evolution of the ambrosia fungus gardening association. Adoption of the fungus-gardening habit allows ambrosia beetles to adopt a ‘generalist strategy’ because the digestive abilities of the fungus widen the range of tree types they can use, wood boring by adults transports the fungi deep within trees, and fungal gardening by the beetle enhances their joint ability to use the biomass-rich resource that the forest trees represent (Farrell et al., 2001).
Ophiostomatoid fungi emerged about 200 million years ago, soon after the appearance of conifers, which are the principal hosts of pathogenic Ophiostoma. This considerably predates the earliest possible origin of the ambrosia beetle-fungus association, 50 million years ago. Raffaelea and Ambrosiella, the two genera of ambrosia fungi, are both polyphyletic and each arose at least five times from the ophiostomatoid clade that includes the obligate pathogens Ophiostoma and Ceratocystis. Phylogenetic analysis estimated the oldest origin of fungus farming near to 50 million years ago, long after the origin of the Scolytinae subfamily (100-120 million years ago). Younger origins were dated to 21 million years ago. Origins of fungus farming corresponded mainly with two periods of global warming in the Cenozoic era, which were characterised by broadly distributed tropical forests. Hence, it seems likely that warm climates and expanding tropical angiosperm forests played critical roles in the successful radiation of different fungus farming groups (Jordal & Cognato, 2012).
Today, invasions of non-native bark and ambrosia beetles are a threat to forests worldwide, and the climatic impact implied by the evolutionary story told in the previous paragraph seems likely to be played out again. Study of invasion patterns in the USA reveal differences between bark beetles and ambrosia beetles depending on their differing ecology. Bark beetles are less dependent on climate, which allows them to colonise more areas within the USA, while non-native ambrosia beetles, are dependent on higher rainfall and warmer temperatures (Rassati et al., 2016). It is the sudden appearance of pathogenicity in insect-fungus symbioses that is the new and currently uncontrollable threat to forest ecosystems, as well as fruit and timber industries, around the globe. Increasingly, perhaps triggered by climate change, some invasive bark and ambrosia beetle/fungus symbioses are shifting from non-pathogenic saprotrophy in their native ranges to a prolific tree-killing in invaded ranges; new and significant diseases are emerging in the world’s forests (Hulcr & Dunn, 2011; Keskitalo et al., 2018; Pasanen et al., 2018).
Updated July, 2019