
12.2 The formal terminology of developmental biology
Development is formally defined as the process of change and growth within an organism during the transition from embryo to adult. This definition immediately creates a major challenge for any mycologist interested in development, because it specifies ‘embryo’, an inevitable result of the fact that most developmental biologists deal with animal systems. Fungi do not have embryos. In fact, fungi are modular organisms in which the multicellular structures (especially fungal fruit bodies like mushrooms, etc.) are appendages of their mycelium, they are NOT individual organisms. This is a feature that fungi share with clonal corals and vegetatively-propagated plants and algae (Harper et al., 1986; Andrews, 1995; Monro & Poore, 2004). The most succinct description of the modular growth form as it applies to fungi (Andrews, 1995) notes that modular organisms are:
‘…characterised by an iterative, indeterminate mode of growth; internal age structure; localised rather than generalised senescence; cellular totipotency; and passive rather than active mobility.’
If you think about it, that quotation describes the fungal mycelium, which features numerous instances of the same hyphal forms extending into new territory for an unstipulated time. The mycelium being comprised of hyphae of all ages; youngest (newly formed) hyphal apices at the growing margin with maturing hyphae towards the ‘older’ more central parts of the colony, and perhaps senescing and dying hyphae at the very centre. Yet even from that senescent centre, inocula can be transferred to fresh medium with the expectation that ‘cellular totipotency’ will ensure that new hyphal tips will be formed from even the senescent hyphae. And, of course, the fungi have an absorptive, sessile lifestyle, so they are not as mobile as any animal, but the exploratory hyphae do invade new territory. Andrews (1995) goes on to point out that:
‘…differentiation of a few basic cell types gives rise to multicellular complexes associated with foraging, reproductive, survival, or dispersal activities.’ And finally: ‘…morphological plasticity of fungi is analogous to but surpasses that of other modular creatures such as the benthic invertebrates and plants. For instance, in response to environmental signals, fungi can vary the timing, extent, and mode of differentiation; interconvert among different growth forms; and decouple the sexual and asexual phases of the life cycle… (Andrews, 1995)’.
So, if the lifestyle of fungi is so different from that of the metazoa used to establish the basic principles of developmental biology, where do we go from here? Well, we suggest a ‘pick-and-mix’ approach. Let’s compare fungal development with animal development and borrow those aspects of the concepts that apply to animals that suit our purposes.
Early development of something like a mushroom looks much like an embryonic process (Fig. 1), and it is clear from images like this that we gain a lot by being able to use the vocabulary of animal embryology, For example: the series of images shown in Fig. 1 clearly show development of the basic body plan of a mushroom; the pattern formation that demarcates the main tissues and organs obviously takes place at a very early stage because the veil, cap, gills, stem and stem base are all clearly established in a tiny structure barely 1% the size of the final mature mushroom (just like an embryo!). We say that we have to use the established language of developmental biology, but we have to apply it with care, understanding just how different from each other plants, animals and fungi are in matters that influence multicellular development.
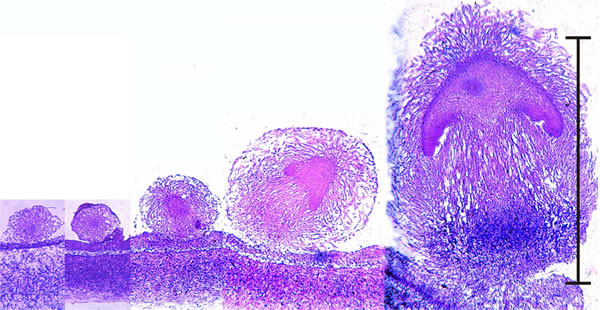 |
| Fig. 1. Early development of something like a mushroom looks very much like an embryonic process (scale bar = 1 mm), but remember that fungi are clonal organisms and these objects are fruit bodies, and many of them may be produced by an individual mycelium over an extended period of time. These images are photomicrographs of light microscope sections of successive stages in the very early development of the fruit body of the Ink Cap mushroom, Coprinopsis cinerea. The mature fruit body is approximately 100 mm tall, so this sequence covers just the first 1% of its developmental programme. The sections have been stained with the Periodic Acid-Schiff reagent, which stains polysaccharide accumulations blue-purple; in this case the polysaccharide, identified by other analyses, is glycogen. The object at extreme left is a large hyphal tuft, at second left is an initial (it shows some internal compaction and differentiation and can become either a sclerotium or a fruit body, depending on environmental conditions). Note that the third section is obviously differentiated into cap-like and stem-like structures, even though it is only 300 μm tall, and this is even more evident in the fourth section (700 μm tall), which has young gills but no gill cavity. At extreme right is a 1.2 mm tall fruit body primordium, in which the basic ‘body plan’ of the mushroom (see Fig. 2) is complete with clear demarcation into veil, pileipellis (= cap epidermis), cap, gills (with the beginnings of an annular gill cavity), and stem (with a distinct stem basal bulb which features heavy accumulations of glycogen). |
Remember that the main eukaryotic kingdoms are thought to have separated in evolution at some single-cell level (discussed in Chapter 2, CLICK HERE to view), so the three eukaryote groups that make multicellular structures became distinct from one another long before the multicellular grade of organisation was established in any of them and must have evolved the mechanisms and mechanics of multicellularity quite independently. There is no reason to expect that the mechanisms that have developed will have anything in common. They will, of course, share all those features that clearly categorise them as eukaryotes, but there is no logical reason to expect that these three Kingdoms will share any aspect of their multicellular developmental biology. If evolutionary separation between the major Kingdoms occurred at a stage prior to the multicellular grade of organisation, then these Kingdoms must have ‘learned’ how to organise populations of cells independently.
The fungal hypha differs in so many important respects from animal and plant cells that significant differences in the way cells interact in the construction of organised tissues must be expected. Inevitably, in many cases these very different organisms needed to solve the same sorts of morphogenetic control problems and evolution may have converged on some common strategies but the outcomes will be analogous (in the evolutionary sense, this mean similarity in function and/or structure between organs of different evolutionary origin), rather than homologous (a similarity of form or structure due to common descent.).
For the most part, though, the many differences in cell biology between animals, plants and fungi result in very different emphases in the way these groups manage their developmental biology. Even in lower metazoans, a key feature of embryo development is the movement of cells and cell populations to remodel an initially unstructured group of cells (Leptin, 2005; Williams & Solnica-Krezel, 2017); evidently, cell migration (and everything that controls it) must play a central role in animal morphogenesis. In contrast, plant cells are encased in walls and have little scope for movement, so their changes in shape and form depend on control of the orientation and position of the mitotic division spindle and, consequently, the orientation and position of the daughter cell wall which forms at the spindle equator (ten Hove et al., 2015).
Fungi are also encased in walls, of course; but two peculiarities, which we have mentioned in earlier Chapters, of their basic structural unit, the hypha, result in fungal morphogenesis being totally different from plant morphogenesis. These are that:
- a hypha grows only at its apex (CLICK HERE to view the section on Mycelium growth kinetics in Chapter 4 and CLICK HERE to view Cell biology of the hyphal apex in Chapter 5); and that
- cross walls form only at right angles to the long axis of the hypha (CLICK HERE to view Septation in Chapter 4) (Harris, 2001).
The consequence of these features is that no amount of cross wall formation (‘cell division’) in fungi will turn one hypha into two hyphae (Field et al., 1999; Momany, 2001; Steinberg et al., 2017). The crucial understanding that makes fungal developmental morphogenesis distinct from both animals and plants, is that fungal morphogenesis depends on the placement of hyphal branches. To proliferate a hypha must branch, and to form the organised structure of a tissue the new hyphal apices must be formed to initiate branches and the position at which the branch emerges from its parent hypha and its direction of growth must be precisely controlled.
Study of multicellular development in fungi (the true analogue of animal embryology) has mostly been done as part of taxonomic studies and has made no contribution to theoretical developmental biology. This is surprising given that so much of what we know about eukaryotic biochemistry, molecular cell biology, cell structure and the cell cycle derives from work with yeast. Still, we can use the concepts and vocabularies arising from developmental study of animals and plants to show how fungi differ.
The word morphogenesis is generally used to encompass the development of the body form of an organism. It has been used to describe the yeast-mycelial transition in some fungi, but this is an aspect of cell differentiation rather than morphogenesis. In animals and plants the key problems in morphogenesis have always related to the interactions between cells from which the patternings of cell populations arise and from which the morphology of the embryo emerges, and the challenge is to apply this sort of approach to fungi.
Comparison of the way similar functions are controlled in different organisms can reveal whether and how different cellular mechanisms have been used to solve common developmental demands (Meyerowitz, 1999). Although, of course, fungi do not feature in such discussions. We intend to feature them, and we will do this by showing some of the experiments and observations that have led to the understanding we currently have of fungal multicellular development. This will involve delving into some of the older literature and will also show how simple experiments and quantitative observations can contribute to important inferences about the fundamental processes involved in fungal developmental biology. But don’t worry; when we have a sufficiently sound foundation, we’ll also deal with the molecular biology.
Updated July, 2019