
11.2 Fungi in food webs
Many of the saprotrophic Basidiomycota that are involved in plant litter recycling, particularly in woodland, form extensive mycelial networks at the soil-litter interface. At the growing fronts individual hyphae are evident in these mycelia, but elsewhere the hyphae often aggregate into strands and cords specialised for translocation of water and nutrients between locations. We have described this before (Chapter 9, in the section titled Linear structures: strands, cords, rhizomorphs and stipes), but here we want to emphasise the size of this mycelial network both in terms of the quantity of mycelial biomass and its physical extent over the ground. This is where we venture into the garden. In the Autumn of 2006 a troop of Clitocybe nebularis (common name: Clouded Agaric or Clouded Funnel) appeared in a secluded suburban garden in Stockport, Cheshire, UK (Fig. 2).
 |
Fig. 2. A troop of Clitocybe nebularis fruit
bodies in a suburban garden in Stockport, Autumn 2006. Observations
began on October 21 and continued for 29 days to November 19. Fruit
bodies of Coprinellus micaceus emerged, matured and decayed
around October 26 and November 1 (one such instance arrowed on the left
above). At bottom right note the corner of a paving slab which was
raised to reveal the mycelium shown in Fig. 11.3. Photograph by David
Moore. |
Observations began on October 21 and continued for 29 days to November 19. At the start, the fruit bodies were young but close to maturity (5 cm diameter), Fig. 2 shows the troop on October 29. The first overnight frosts of the season occurred on October 31 and November 1; temperatures were otherwise generally above average throughout October in that year. Some of the mushrooms were close to a paved area and when one of the paving slabs was lifted the mycelium that had penetrated beneath it was evident (Fig. 3, photographed, as were other mycelium images, on November 1).
 |
Fig. 3. Removal of the paving slab revealed the
extensive mycelium of Clitocybe nebularis growing beneath it.
Numerals show the locations of Figs 11.4 to 11.8. Photograph by David
Moore. |
 |
|
|
 |
Figs 11.5 to 11.8. Lush and extensive hyphal growth of Clitocybe nebularis beneath the paving slab, with mycelial strands emerging from litter fragments. Positional key for these images shown in Fig. 3. Photographs by David Moore. |
Mycelium was evident throughout both soil and litter layer (Fig. 4), and mycelium beneath the 60 cm square paving slab covered soil particles, leaves and dead and dying roots with a lush growth of fine hyphae (Figs 5, 6) that aggregated into strands and cords where it colonised the more discrete, and patchily distributed, debris such as root fragments and larger leaves (Figs 5, 7, 8). As this mycelium is supporting growth of the fruit bodies shown in Fig. 2, it is quite clear that it must persist for several days to a few weeks. Yet it is equally clear that the biomass making up such a mycelium must be a tempting food resource for a range of organisms.
Conventional ideas about nutrient cycling in the soil food web assume that bacteria consume easily broken-down substrates faster than fungi, and that mycorrhizal fungi do not decompose organic matter; these views are now being challenged. However, considerable evidence has emerged showing that saprotrophic fungi consume significant quantities of the soil’s readily available nutrients, and that mycorrhizal fungi decompose organic matter to obtain carbon, nitrogen, phosphorus and sulfur for their own and their host’s needs (see Chapter 13). Regarding available carbon, De Vries & Caruso (2016) suggested a new conceptual model for the lowest trophic level of the soil food web, in which organic carbon consists of a continuous pool (rather than different pools labelled ‘labile’ and ‘recalcitrant’), with saprotrophic fungi using substantial amounts of labile nutrients in a balanced coexistence with bacterial saprobes, together with decomposition of organic matter by mycorrhizal fungi.
In natural woodland large and persistent mycelial networks like those illustrated in Figs 11.5 to 11.8 are common and their ability to recycle soil nutrients results in them being a food resource that is the foundation of several important food webs. There have been numerous studies on interactions between fungi and soil invertebrates but most of the scientific studies done so far have concentrated on grazing of fungal mycelia by microarthropods known as collembola (Tordoff et al., 2006, 2008; Wood et al., 2006). Collembola are integral components of the soil environment, feeding on many organisms including bacteria, lichens and decomposing material but they do prefer fungal hyphae over all other food sources.
Collembola (also known as springtails) are primitive wingless insects that range in size from 0.2 mm to 9 mm. They are plentiful in all soils and population densities commonly reach 104 m-2 so hyphal grazing by these invertebrates can result in dramatic changes to mycelial morphology. They express intriguing food preferences in laboratory experiments (Rotheray et al., 2009). Collembola such as Folsomia candida graze the older parts of the mycelium but appear to prefer hyphal tips and fine mycelium; other collembola choose to feed on the hyphae of conidial fungi rather than hyphae of arbuscular mycorrhizal fungi. Grazing on this scale can impede the spread of mycelial networks through woodland and their ability to decompose dead organic material (Tordoff et al. 2006, 2008; Wood et al. 2006). In response to grazing damage the mycelium attempts to evade the attacking fungivore by diverting energy into accelerated growth away from the grazing animal, and in a mycorrhizal mycelium this can result in enhanced exploratory growth of young hyphae and improved nutrient supply to the host plant (Ngosong et al., 2014). Aside from contributing to food webs through predation, the main environmental impact of the collembola is to mineralise nitrogenous and phosphorus-compounds by digestion and excretion into the soil, and this leads to increased plant biomass. This positive effect will, of course, be offset by any damage done to mycorrhizal mycelium although collembola tend to occupy the upper litter layers where saprotrophic fungi predominate.
Mites are at least as numerous in soil as collembola and are also very small, being in the region of 250 μm long. Most are predatory, feeding on nematodes and other arthropods, but there are others that favour bacteria and fungi. Red pepper mites (Pygmephorus spp.) feed only on Ascomycota, particularly Trichoderma spp., and are often associated with composts (including mushroom farm composts) in which these moulds can be abundant. They are not primary pests of the mushroom farm but they indicate the presence of the Trichoderma weed mould, which can itself cause mushroom crop losses. The most important mite pest of mushroom farms is Tarsonemus myceliophagus; the clue is in the name! This is a very small animal (about 180 μm long) that feeds preferentially on fungal mycelium. Because of its small size it can go unnoticed until damage symptoms appear in the mushroom crop (Rinker, 2017).
Many species of nematode occur in habitats rich in decaying organic matter (Glavatska et al., 2017). Some are saprophagous (feeding on decaying materials), others are mycophagous (specifically fungal feeders). These are small colourless worms, up to 1 mm long, that swim in the surface films of water on soil and litter fragments. Two species of mycophagous nematodes that feed exclusively on fungal mycelium are Dictylenchus myceliophagus and Aphelenchoides composticola. Either of these can increase in number by 50 to 100 times in a week so they can rapidly reach population levels sufficient to totally destroy mycelium.
The larvae (or maggots) of many small dipteran flies are probably the most frequently seen mycophagous animals; it is rare to find a mushroom in the field that is completely free of maggots. Adult females lay their eggs on the fungal fruit body and when they hatch the larva eats its way into the tissue, and subsequently eats and tunnels through the fruit body tissue, causing enormous damage (Figs 9, 10). These are ‘mushroom flies’; they are about the size (one to a few mm long) of fruit flies (family Drosophilidae), to which they are related. Indeed there is one fruit fly, Drosophila funebris, which lays its eggs in mushroom cap tissues. However, the specialised mushroom flies belong to the related families Sciaridae (commonly known as dark-winged fungus gnats), Phoridae (hump-backed flies, also known as scuttle flies because of their habit of escaping by running rather than flying) and Cecidomyiidae (called cecids, are gall midges or gall gnats, the larvae of most of which feed in plant tissues, creating abnormal growths called galls). Larvae of sciarids tend to tunnel from the bottom of the mushroom stem upwards, rarely burrowing into the cap; damage caused by phorids tends to be from the top of the mushroom downwards. Both also eat mycelium. Cecid larvae feed on mycelium and the base of primordial mushroom stems; some also have a taste for mushroom gills.
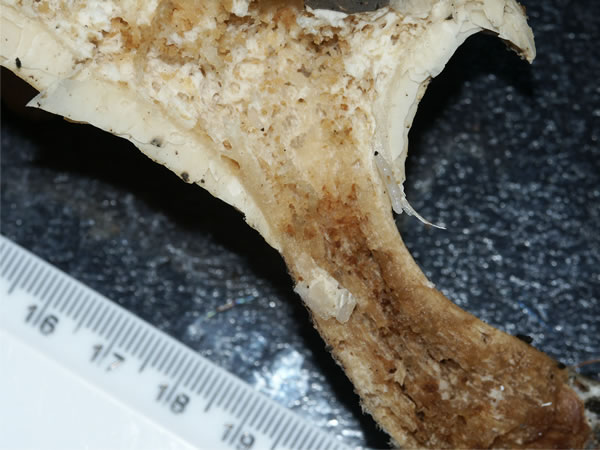 |
Fig. 9. A sectioned fruit body of Clitocybe
nebularis showing extensive tunnelling by larvae (maggots) of
mushroom flies. Photograph by David Moore. |
 |
|
|
There are a few species of sciarids that are common, and one, Lycoriella solani, has become a very damaging pest of mushroom farms because it has developed resistance to organophosphorus insecticides. Adults can affect the crop by spreading mites, bacteria and spores of weed fungi around the mushroom house. Serious crop damage is caused by the larvae tunnelling through the stems of maturing mushrooms, but the most critical damage results from infestation early in crop growth because the larvae can destroy developing mushroom primordia and completely devastate the crop.
Phorid flies are common, their larvae are generally omnivorous, eating anything from decaying vegetation to fresh meat, and these flies are very diverse; the genus Megaselia alone having over 100 species in the UK. But one species, Megaselia halterata, has become an important pest of mushroom farms because the larvae eat mushroom mycelium (the female is attracted to freshly-growing mycelium to lay her eggs) and the adults spread the disease-causing fungus Verticillium. Larvae of other species of Megaselia feed on and in developing mushrooms, causing tunnelling damage to mushroom cap and stem. Megaselia nigra is the species most often found causing tunnelling in wild mushrooms and can become a pest in mushroom farms.
Cecidomyiidae are so small that the adults are seldom seen, so although over 3000 species have been described there are probably more awaiting discovery. Many species are economically important as pests of crop plants. Others produce predaceous larvae that prey on other arthropods. Six species have been found on mushrooms, the most frequently encountered being Heteropeza pygmaea. An unusual feature of the cecids is that the larvae reproduce by paedogenesis, an asexual process in which a cecid larva becomes a ‘mother larva’ from which 10 to 20 daughters emerge within a week of the mother larva hatching from its egg. This method of reproduction leads to rapid multiplication that produces larval swarms capable of doing enormous damage to mycelium and mushrooms (Rinker, 2017).
When the mycelium makes fruit bodies (like mushrooms, brackets and truffles), these are also vital food sources for many animals, from large arthropods and molluscs, to small mammals and humans. Slugs and snails can eat large cavities in the caps and stems of mushrooms, and a large slug can easily consume an entire mushroom (Fig. 11). Many small mammals depend on fungal fruit bodies, especially hypogeous ones like truffles, for a significant part, even the majority, of their diets. This extends the influence of fungi on the food web right up to the top predators, like birds of prey. Those that eat the small mammals that subsist almost entirely on fungal fruit bodies, are themselves indirectly dependent on the fungi of the soil.
 |
Fig. 11. An orange form of the large black slug,
Arion ater, completely demolishing a Coprinellus micaceus
mushroom fruit body. Photographed by David Moore in the same garden in
Stockport. |
A celebrated example of this is the Northern spotted owl (Strix occidentalis caurina), in the Pacific Northwest of the USA. This bird has a wingspan of about 1.2 m and ranks as one of the largest birds in North America. Pacific Northwest communities rely heavily on timber harvested from Federal forests in the area, and for many years when old-growth forest was harvested, the cleared areas were replanted with fast-growing commercial tree species, rather than the original, slow-growing, species. Unfortunately, despite this reforestation the population of the northern spotted owl declined drastically. There are only 500 pairs in the state of Washington, so the death and/or migration of relatively few animals could drive the population level beneath sustainability.
The owls are very territorial and intolerant of habitat disturbance; they prefer old-growth forests with high and open tree canopies so they can fly between and beneath the trees. Their staple diet consists of small mammals: squirrels, wood rats, mice, voles, and most of these small mammals themselves depend on hypogeous fungal fruit bodies (= truffles) for a significant part of their diets. Unfortunately, the fast-growing tree species that were good for commercial reforestation had different mycorrhizas to those of the original forest; they did not produce fruit bodies small mammals would eat. Consequently, populations of small mammals declined, and with the loss of prey species, owl populations also declined. These circumstances contributed to the main lesson about conservation: that to conserve a species it is necessary to conserve its habitat. Harvest of the old growth forest trees could only be made sustainable if cleared areas were replaced with similar communities; using a different range of tree species for commercial purposes produces an entirely different habitat. The Endangered Species Act of 1973 requires the US federal government to identify species threatened with extinction, identify habitats they need to survive, and help protect both. The Northwest Forest Plan of 1994 established these principles as legally binding requirements that helped conserve the owl and its prey species, but went further by listing 234 old growth dependent (and rare) fungal species for additional protection.
A more recent study suggests that the Northern Spotted Owl will require more protection if it is to be conserved (Dugger et al., 2016). During 1985-2013 the survival rates and reproductive outcomes of owls were analysed in 11 areas that represented 9% of the owl’s range across Washington State, Oregon, and northern California. They noted that spotted owl populations declined by 55-77% in the state of Washington, 31-68% in Oregon and 32-55% in California during this time; an average rangewide decline of nearly 4% per year between 1985 and 2013. It appears that the Barred Owl (Strix varia), an invasive species, is out-competing Spotted Owls for space, food and habitat. Barred Owl population densities are high enough across the range of the Northern Spotted Owl that, despite the continued management and conservation of suitable owl habitat on federal lands, the long-term prognosis for the persistence of Northern Spotted Owls may be in doubt (Dugger et al., 2016).
Updated July, 2019