解讀真菌的形態發育
趙紹惠 和 大衛摩亞
Department of Biology, The Chinese
University of Hong Kong, Shatin, N. T., Hong Kong SAR, China
中國香港新界沙田 香港中文大學生物系
&
School of Biological Sciences, University
of Manchester, Manchester M13 9PT, U. K.
英國曼徹斯特市 曼徹斯特大學生物科學學院
第五課:真菌細胞分化和形態發育
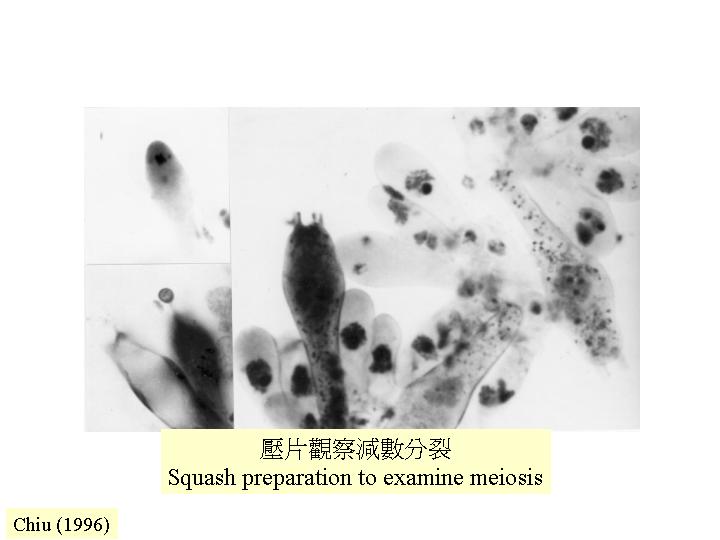
單細胞的分化可閱Chapter 2,這章討論減數分裂細胞、囊狀細胞和子實體的狀況。
單細胞的情況:
1. 減數分裂細胞
(meiocyte)
許多真菌的性器官是多細胞的,需要將器官解剖才可看到減數分裂細胞
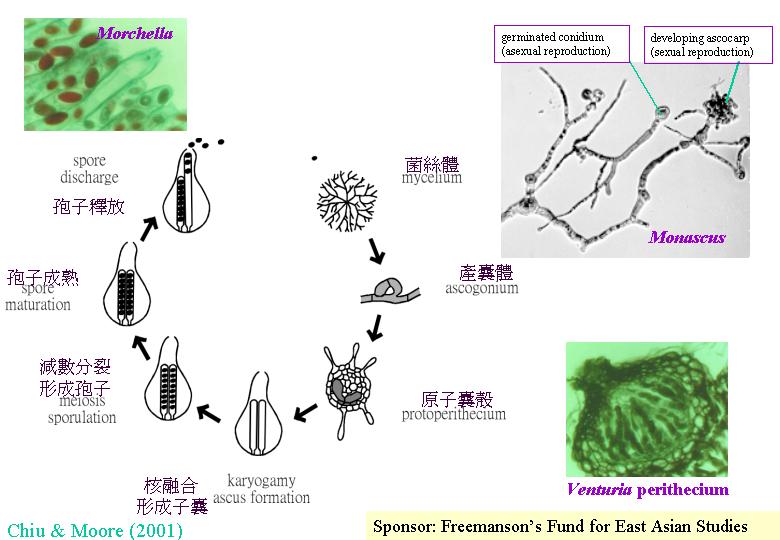
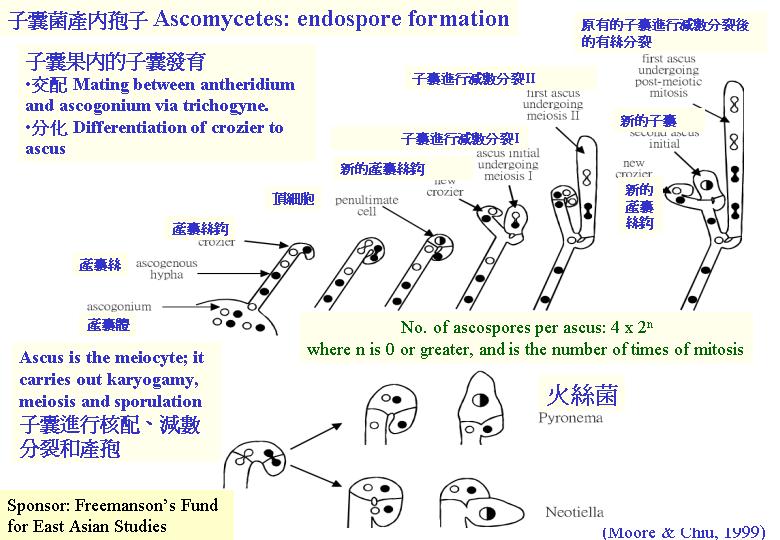
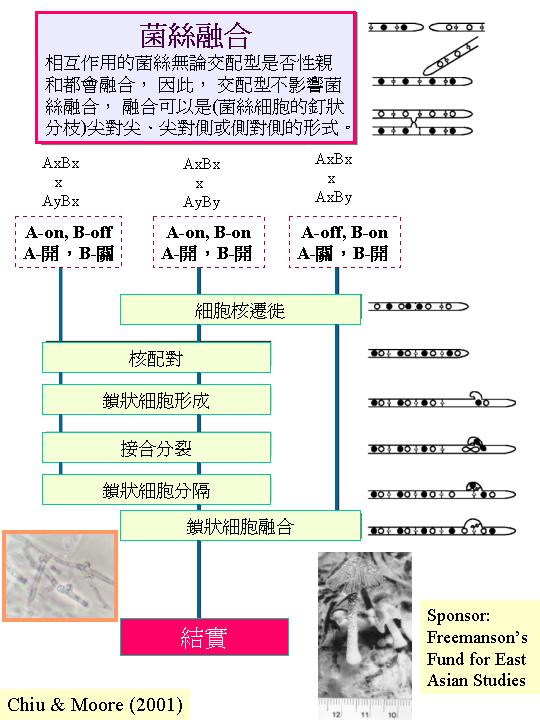
(meiocyte) ,子囊菌的子實體許多都是細小的,只要將它放在玻璃片,製成壓片,便可在顯徵鏡下觀察子囊的形成 (ascomycetes)。雌性和雄性結構間的交配或兩性的菌絲融合導致:雄性細胞核遷移到雌性細胞形成產囊體,當中雌細胞核和雄細胞核配對但不融合(雙核期),產囊體長出產囊絲,大部份細胞保持這雙核狀態(dikaryotic;核配對隨菌絲生長而進行共同分裂 (conjugate division) ,典型的發育情況是:產囊絲彎曲成一個產囊絲(crozier) ,產囊絲內的雙核同時分裂為4個核,和形成2個隔膜,構成3個細胞:產囊絲彎部細胞是雙核而其餘細胞是單核的,雙核細胞發育成子囊母細胞去進行核融合(karyogamy) ,發育成子囊,子囊完成減數分裂產生4個單倍體子核,若再進行有絲分裂,子囊內便有8 個子囊孢子,或更多的子囊孢子( ascus)。
雙倍體釀酒酵母在環境缺乏營養下,會進行減數分裂和產孢,利用操控營養的提供,可將酵母的分化同步化。因此,對這發育過程的專一基因表達 (differential gene expression) 、減數分裂所需的機制 (machinery),和環境因素都可透過研究、觀察而得,再加上基因突變株
(mutants) 的應用,便可確定減數分裂細胞的分化特性。釀酒酵母既然可用簡單的培養基,生長週期短,而操控方便,這方面的結果較容易獲得,相對下,其他真菌的研究便較少。
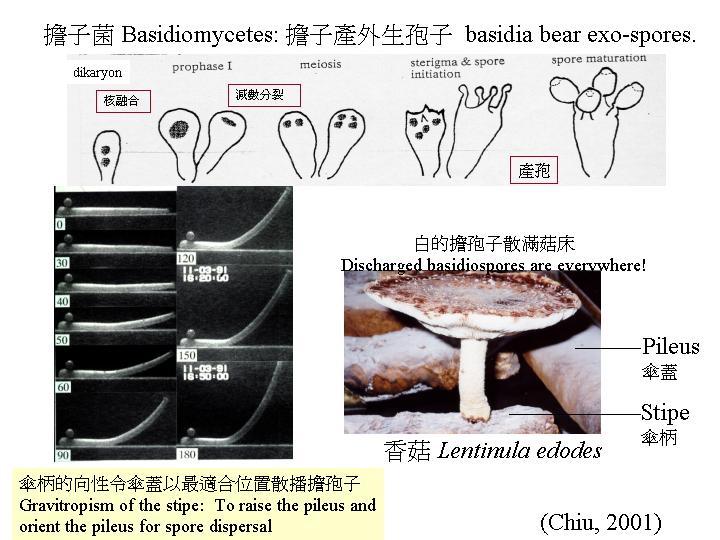
遠在Buller 年代,我們已知道大部份擔子菌的減數分裂是不同步的
(asynchronous) (asynchronous
meiosis; Chiu & Moore, 1999),除了鬼傘屬 (Coprinus species) ,Lu (1982) 看見灰蓋鬼傘 (Coprinus
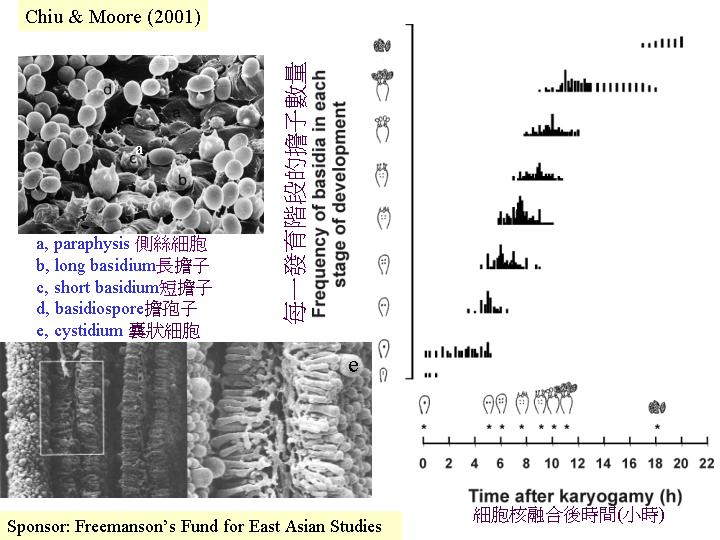
cinereus) 每一時刻有70 –75%性細胞 (擔子) 進行同步的減數分裂,Pukkila et al. (1984) 和Hammad et al. (1993a) 就用子實層看到的減數分裂至孢子成熟的現象,作為一客觀形態發育的指標 (hymenium development)
,這亦廣為應用。
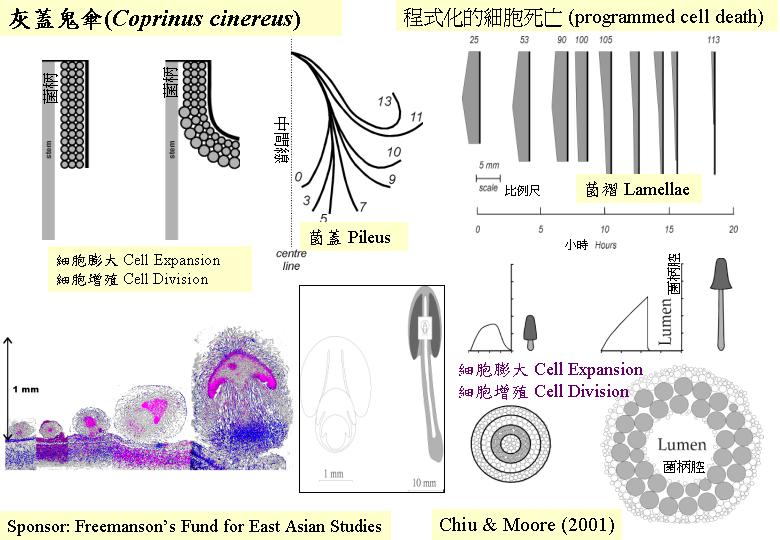
在不同時段,將子實層抽樣製成即看的玻璃片,在光學顯微鏡下,觀察和分類擔子分化的階段,可得關係: 灰蓋鬼傘(Coprinus cinereus) 擔子邁進減數分裂和產擔孢子的時間旅程 (hymenium
development)。0小時代表細胞核開始融合,減數分裂I 在第5 小時發生,減數分裂 II在第 6
小時、擔子梗在第 7.5 小時出現,擔孢子則在第9小時開始出現,第11
小時擔孢子開始深棕色,和第18 小時擔子釋放成熟的擔孢子(Hammad et al., 1993a)。
典型的無隔擔子源於一雙核菌絲的頂端細胞, 這細胞膨大和進行核融合和減數分裂, 在減數分裂的後期, 擔子頂端長出4個擔孢子梗,擔孢子梗支撐膨大生長的擔孢子, (擔子菌生產外生孢子,子囊菌則生產內生孢子) , 擔子內的子核經擔孢子梗進入擔孢子,而且,或會在擔孢子彈射前進行一次有絲分裂
(Chiu & Moore, 1990c; Basidiome)。
在體外移殖環境下, 兩種鬼傘擔子活動速度會放慢 (Bastouill-Descollonges &
Manachere, 1984; Chiu & Moore, 1988a), 正如McLaughlin (1982)用電池場影響在液體栽培的擔子形成擔孢子梗一樣, 擔子由減數分裂前期 I到擔孢子梗形成需要1天時間, Raju & Lu (1970) 測定在子實體上的擔子需要11.5小時完成減數分裂, 另外8-10小時產孢子,灰蓋鬼傘株Meathop只需要10小時完成減數分裂。就算培養基是水瓊脂, 結果都一樣,因此,減數分裂是自主進行的。
發育專一性 (Commitment)
大部份的真菌組織受到騷擾和被接種到一培養基上, 都會長出營養菌絲, 減數分裂期的體外菌褶則會完成產孢子的程序
(Descollonges & Manachere, 1984; Chiu & Moore, 1988a), 所以, 擔子是在減數分裂前期 I 時決定進入產孢子的, 這可引証為真菌發育的專一性。Chiu & Moore (1988a) 的實驗手法被推廣成為一快速、小規模的生物測試法, 這樣地可測試添加的外間物是否調控物質
(modulator), Chiu &
Moore (1988b) 証明胺鹽基ammonium ion 和谷氨酸阻止擔子分化, 擔子停止產孢子而營養菌絲在擔子生長點長出來。在測試pH和氨 (硫酸和氯離子)濃度影響時, 發現:
pH 值為6-8時, 體外栽培的菌褶繼續發育, 高pH值和氨離子的最小抑制劑量達50mM便抑制菌褶發育, 硫酸鉀和氯化鉀卻沒有這效果, 同樣地, 利用微針將20 μl 的
1M濃度氨離子注射到年幼的鬼傘菌蓋內, 會局部阻止發育,形成黑菌褶上的白區(鬼傘生產黑擔孢子, 因而成熟期的菌褶是黑色的), 雙核期的子實體則抵受不了實驗操作的傷害而流產, 其他期間, 若取代以水或緩液注射, 並無影響, 將體外和體內測試的結果比較, 不難發現: 氨離子在體外的環境下, 完全抑制抽離的小片菌褶,但在整個子實體的體內測試時, 只是拖慢減數分裂速度,沒壓抑產孢子。當菌褶到達產孢子階段, 兩種測試法都表達抑制作用
(Chiu & Moore, 1988a) ; 雙核時期的抽離不導致營養菌絲出現;細胞核融合後, 擔子發育受抑制, 但氨離子的作用不只是停止分化, 部份擔子改到營養生長, 因此, 氨離子的直接作用是打破擔子分化的專一性。 而且菌絲就在生長點如:擔孢子梗長出;如擔孢子梗已長出, 取代擔孢子的是菌絲;若擔孢子已出現, 氨離子就不讓它完成發育, 而改成菌絲尖 (ammonium toxicity)
;還有, 菌絲可在擔孢子的底部出現。
Chiu & Moore (1990) 利用同一生化測試法, 考查金屬離子、去膜極化劑、離子戴體, 外加的cAMP和抑制細胞壁合成物,結果是: 氨離子、谷氨胺 和金屬對第一減數分裂後的擔子同樣有效, 而離子戴體、cAMP 和細胞壁合成抑制物對減數分裂的擔子有抑制作用, 因此,產孢子的準備是在減數分裂時做的, 另外, 除細胞核分裂外, 擔子的其他反應如合成細胞壁等, 受考查的金屬離子、 氨離子和谷氨胺影響,這些抑制物都引起由擔孢子梗的位置長出菌絲,因此, 這些位置在分化早期便固定下來的。另外, 許多物質由底物類似物、細胞壁抑制物 (如: nikkomycin) 以至無機鹽如NaNO3
等都引起擔子返回營養生長: 在擔孢子梗位置長出菌絲。將所有相關的實驗作總結, 明顯地有一類: 氨離子和谷氨胺及它的類似物、 無機正離子和負離子等阻止第一減數分裂後的擔子完成分化和引致擔子長出菌絲, 第二類包括離子戴體、cAMP和細胞壁抑制物, 只有對擔子的減數分裂有抑制作用, 而且減數分裂和產孢子兩過程是可分開的。 致於氨離子的反應, 就表明受影響的過程可能與減數分裂無關, 或在產孢子期出現。
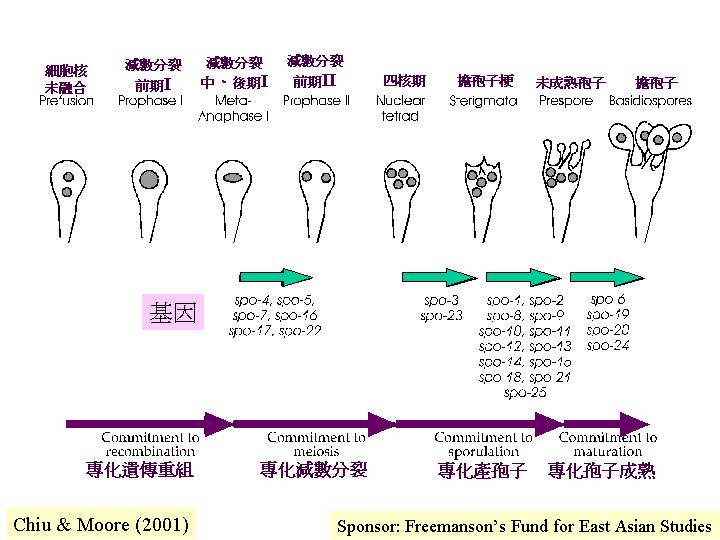
Lu (1972) 提出減數分裂前期 I
前10 個小時就是擔子程式化地進行細胞核融合和染色體配對, Raudaskoski
& Lu (1980) 就用hydroxyurea 去抑制DNA合成, 結局是擔子停在雙核期, Lu (1982) 提出這是決定性階段: 擔子繼續分化進行遺傳重組 (genetic recombination) 。套用研究釀酒酵母時常用的詞匯, 這分化的途徑就是: 專一化作遺傳組合,這不等同專一化作減數分裂, 而後者亦不等同專一化作生產擔孢子
(Berry, 1983; Dawes, 1983) 。
Raju & Lu (1973) 發現鬼傘擔子在雙線期 (diplotene)
時,已進行合成紡錘體 (spindle
pole body) , 這應該是分化的檢查點
(checking points) , 灰蓋鬼傘和釀酒酵母呈現共同的要求和出現遺傳重組和減數分裂兩個分化檢查點。 不同處是:若在這兩個檢查點期間, 將釀酒酵母抽離產孢培養基, 細胞會返回營養的生產模式(Berry, 1983), 但在雙核期抽離灰蓋鬼傘, 擔子是停止發育的, 其他組織如側絲細胞會返回營養式的生產模式,掃描電子顯微鏡可見到這些新生菌絲甚至穿插到子實層面去(ammonium
toxicity;Chiu & Moore, 1988) 。
而減數分裂細胞 (meiocyte) 的分化有幾個里程碑/
檢查點 (Basidial
differentiation )。
a. 對基因重組的分化專一性 (要求完成DNA的合成, Lu, 1982) ,
b. 對減數分裂的分化專一性 (於前期一(prophase
I)階段, Lu &
Chiu, 1978; Chiu & Moore, 1988a) ,
c. 對產孢子的分化專一性 (於第二減數分裂後,
Raudaskoski & Lu, 1980; Chiu & Moore, 1988a, b) ,
d.
對擔孢子成熟的分化專一性 (Chiu & Moore, 1988b) 。
有趣的是:釀酒酵母 (Saccharomyces
cerevisiae) 的氨透性 (ammonium
permease) 探測氨離子和激活 cAMP訊息路徑,提升的cAMP水平致使假菌絲
(filamentation (formation of pseudohyphae)) 的出現
(Lorenz & Heitman, 1998)。
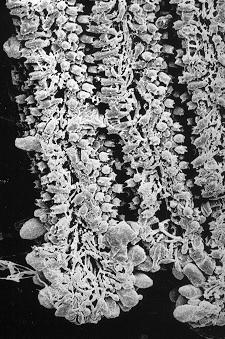
若從子實層構造看, 分佈廣而獨立的囊狀細胞、接囊狀細胞和子實層主要細胞類:側絲細胞 (Coprinus hymenium)
都沒有專一的分化, 只有擔子才是一細胞分化模式, 正因如此, 正常的子實層發育, 就需抑壓側絲細胞的生長, 好讓系統模式表現出來。
2. 囊狀細胞 (Cystidia)
擔子功能是細胞核融合、減數分裂和形成擔孢子這一命運途徑,子實層上可找到其他類的細胞,如:灰蓋鬼傘 (Coprinus cinereus) 的子實層有囊狀細胞
(cystidia) 和側絲細胞 (paraphyses)
。長久以來,囊狀細胞的形態多被用作分類用途, 囊狀細胞都被認為是不育而繼續生長的擔子 (Brefield, 1877; Corner, 1947) ,其實, 囊狀細胞的命運與擔子的不同, 它有自己的特定功能,而許多研究都忽略了形態發展的彈性
(developmental plasticity) 。
灰蓋鬼傘的囊狀細胞都從菌褶髓的菌絲長出(Moore et al., 1979;
Rosin & Moore, 1985b), 所以, 過程包括分化
(cell differentiation) 和固定發展途徑。最初的子實層有擔子和8%
是體積較大的囊狀細胞 (Horner
& Moore, 1987),而裂褶菌 (Schizophyllum
commune) 的擔子則是由子實層子層
(subhymenium)鎖狀連合生長出來的 (Niederpruem
et al., 1971; Niederpruem & Jersild, 1972) , 成熟的鬼傘子實層還有側絲細胞 (paraphysis)
, 它們的膨大就隔開成長中的擔子
(Moore et al., 1979), 約75%側絲細胞在減數分裂時出現
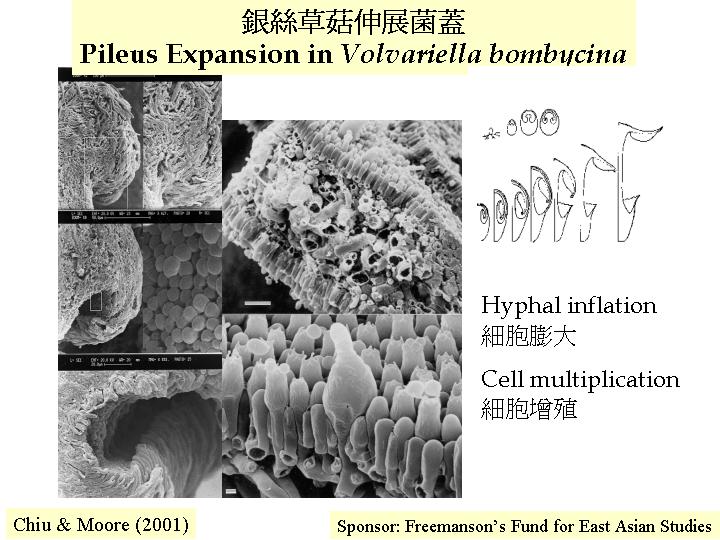
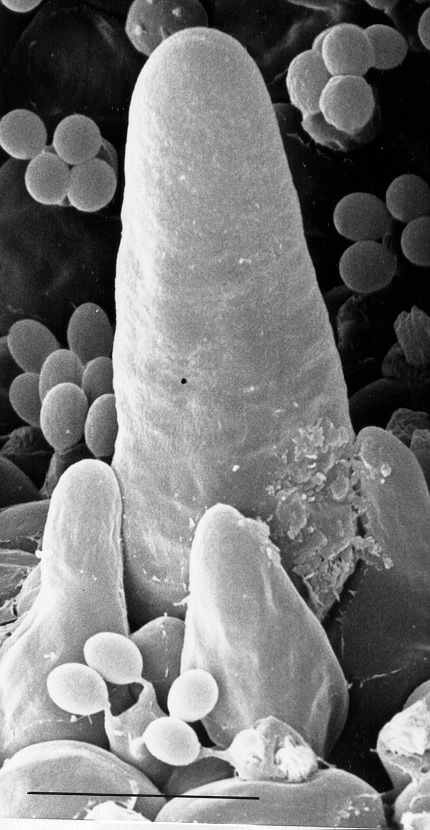
(Rosin & Moore, 1985b), 其生長模式是:1-2 mm的子實體原基內的子實層是一層緊密的年青擔子和較大的囊狀細胞,而隔絲細胞剛在子實下層形成並未穿插出現在子實層。中期,隔絲細胞先長至擔子的2/3高度時,不延伸,只膨大 (圓而不定向的生長) ,隔絲細胞成長、膨大和穿插出現在子實層去分隔擔子,而隔絲細胞與擔子在子實下層仍是相連的。到最後, 一個擔子由五個側絲細胞包圍, 即80%以上的子實層細胞是作機械性支撐功能的, 另外,銀絲草菇與鬼傘一樣在菌褶邊和平面上都有囊狀細胞。
灰蓋鬼傘的面囊狀細胞 (facial cystidia) 由子實子層生長出來, 因而是雙核的, 邊囊狀細胞
(marginal cystidia) 就源自多細胞核的菌褶髓, 因而也是多細胞核的 (Chiu & Moore, 1993), 銀絲草菇的囊狀細胞則都是多細胞核的,而囊狀細胞有水珠凝結, 可推斷為分泌功能 (Chiu & Moore, 1990b), 但鬼傘的面囊狀細胞有建築工程的功能:
Buller (1909)和 Chow (1934)估計面和邊的囊狀細胞將兩面對的菌褶隔開 (facial
cystidium), 防菌褶相撞, 但情況並非如此;
一: 鬼傘菌褶的菌髓是一開放組織和充滿空位 , 沒有牢固的基底,
二:囊狀細胞橫跨兩菌褶,與對面子實層的連囊狀細胞(cystesia)
牢牢連結,calcofluor
white螢光劑的染色顯示:嶄新合成細胞壁上的幾丁質位於囊狀細胞與連囊狀細胞連結處(Chiu
& Moore, 1990),相對下,囊狀細胞與連囊狀細胞強化隔鄰子實層的連結。
若作為分隔, 這連結就不需要, 固此, 面囊狀細胞功能是作為兩菌褶的橋樑,連接鄰近的菌褶成為一子實層體
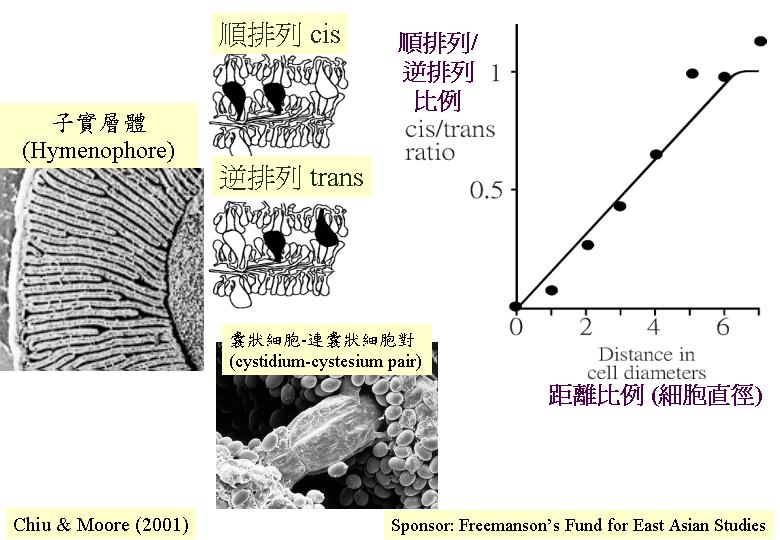
(hymenophore), 當菌蓋擴張時, 其拉力便透過囊狀細胞-連囊狀細胞對(cystidium-cystesium
pair)來平均分配到整個子實層體去。
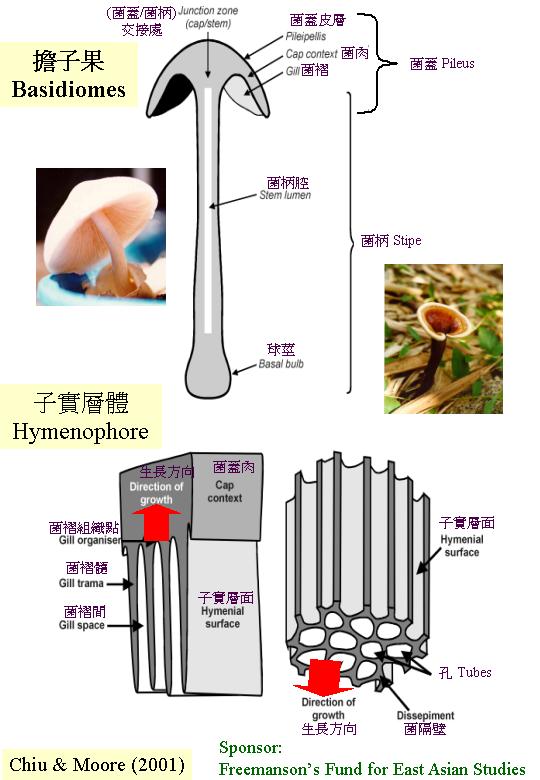
多細胞的性器官- 子實體
1. 多細胞的性器
真菌組織的基本結構稱「密絲組織」(plectenchyma), 其詞根是:希臘文 plekein(意謂編織)和enchyma (聯合), 因此, 詞匯生動地表達真菌菌絲藉互纏而形成的結構。基本的「密絲組織」有二類型:疏絲長軸組織 (prosenchyma;希臘文 pros即向、近似的意思), 專指一鬆弛互纏的菌絲組織;和擬薄壁組織 (pseudoparenchyma;希臘文 pseudo 即假、非真確的植物薄壁組織)。
Butler (1958) 、Townsend & Willetts (1954) 和 Reijnders (1977, 1993) 在許多不同種屬的球基、菌柄、菌幕、傘蓋、菌肉、菌絲索 (cord) 、菌絲線 (strands)
、菌核 (sclerotia) 等可見到菌絲扭結, 此為一團互纏協調分化的菌絲, 有些像是休眠, 有些已分化: 中央菌絲由一環或圓柱體的細胞包圍。菌絲扭結可能是真菌最基本的調控形態分化的目標物 (morphogenesis)。菌絲線是遷移的器官,負責去尋找食物源。 干朽菌 (乾腐爛菌Serpula lacrimans) 的菌絲索 (mycelial cord) 可穿透幾公尺的砌磚來覓食木材、甚至塑膠薄膜和建築材料,靈芝就生產假菌絲線,內有底物如泥粒、碎石和菌絲(Cheung & Chiu,
unpublished result)。
相對菌絲線,菌索 (rhizomorph) 表現嚴謹的、如植物根的分化和組織。樹和灌木的病源菌
(pathogen) 蜜環菌(Armillaria
mellea),當中有許多形態近似的不同種;speices
complex) 的菌索將森林中樹木根部連接(Rishbeth,1985)。菌索負責雙向的運輸和遷移的功能(Granlund et al., 1985)。菌索擁有一高度組織的前端生長點而且展現極端的頂優勢 (apical
dominance)。 在菌索的前端區域有緊密的細胞相連, 然後接連是中髓部份, 散佈於這區域的空間或黏液上有肥大的、具液胞的和通常是多核的細胞, 這個中髓區域形成菌索的中央通道。 成熟的菌索中髓部份縱橫交錯著狹窄的纖維菌絲和寬直徑的導管菌絲, 外皮層的細胞則較小、較黑, 而且有厚壁, 甚至外延有擴散的菌絲,與植物根相似。菌索直切面展示前端為緊密排列的細胞,菌索髓區則有肥大的、有空泡的和常是多細胞核的細胞,細胞間則充滿空氣或黏液,除此外,髓區的中央通道有狹窄的纖維狀菌絲和成熟的寬直徑管菌絲。
另外,潮濕森林中暴露於空氣的菌索主要是由小皮傘 (Marasmius
spp.) 架設的, 將墮落的鮮葉攔截, 形成懸掛的垃圾層(Hedger, 1985; Hedger et al., 1993)。 Hedger et al. (1993) 發現這些菌索尖有一退化似的菌蓋保護免受乾燥。這些空中的菌索與泥土中的功能是類似的, 但發育上則與菌柄的不定生長類似 (Jacques-Felix, 1967)。
許多學者推斷菌索有分生組織 (meristem),如:擔子果體內可找到平行而快速生長的組織層。Reijnders (1977) 就時常將擔子果分層定界線,並稱為 meristemoids (類分生組織層)。植物分生組織的細胞在各個不同的平面都可進行細胞有絲分裂, 產生一有秩序的組織, 形成整個頂端區域一起生長,其他部分則負責分化或特化的功能。這對真菌是絕對錯誤的。
形態發育可視為一至數個平行而獨立的子程序(subroutines) (development)
。Chiu et al. (1989) 認為操控和協調擔子果形態發育這些子程序 (有形的結構如:
底座、菌、孢子等, 和「無形」的操控), 便成為標準生長發育的模式, 這個想法和 Gregory (1984) 和 Rayner & Coates (1987) 營養菌絲的「模態開關」(mode switch) 概念是一致的。但是,協調子程序的工作若沒落實,畸形/異形發育使出現;若複制菌褶過程不停止和在擔子果成熟期繼續出現, 則子實層體會出現複雜如蜜蜂巢的情況, 與Lentinus 香菇屬或銀絲草菇的超多子實層體 (supernumerary hymenia) 發育異種所見的一樣 (Chiu et al., 1989; Hibbett et al.,
1993a)。
最早期的真菌分類源於Fries (1821), 是因循Persoon
(1801)的資料將各品種分類,同一進化來源但不同演變(homology)、不同演化但結果類像(analogy) 等的進化概念在真菌學界很晚才出現。擔子果分類的考慮條件包括: 產孢子組織的形狀和形式, 子實層和子實層體構造, 傘菌指子實層體以菌褶在菌蓋下方存在, 多孔菌的子實層體以孔存在 (如牛肝菌和支架菌), 而裸露菌 (hydnoids)其子實層體為齒或刺狀, 裸露菌的擔子果是平放在底物上 (resuspinate), 其他形式如:權頭狀的(clavarioid), 珊瑚狀的
(coralloid), 還有, 擔子果不開放的 (gastroid)。
Watling (1978, 1985, 1996) 介紹和描繪各種情況, 指出有超過十種擔子果形, 這些都同能在外露環境下保護子實層: 裸果gymnocarpic到各種不同形式, 菌幕將菌絲包圍子實層, 如要調控這封閉的環境,可靠操控囊狀細胞的分泌 , 因此, 菌幕是有保護作用的。
擔子果有不同構造 (basidiome
polymorphism):
i.
裸果型的發育模式 (gymncarpic
development) :子實層從開始到成熟就裸露。
ii. 四類型的菌幕被果型的發育模式
(velangiocarpic development) :子實層由菌幕包圍保護,直至擔子果成熟釋放擔孢子時。
·
單一菌幕被果型 (monovelangiocarpy): 原基被外菌幕 (universal veil) 包圍,
·
雙菌幕被果型
(bivelangiocarpy):除外菌幕外,子實層被內菌幕
(partial veil)包圍,
·
副菌幕被果型 (paravelangiocarpy):僅有的內菌幕成熟時會退化消失,
·
異菌幕被果型
(metavelangiocarpy):菌傘和/或菌柄衍生類內菌幕組織。
iii. 菌幕裸果型的發育模式 (gymnovelangiocarpic development) :子實層只在初期由一極退化的菌包圍。
iv. 蓋緣成長的被果型發育模式 (名蓋生被果型,pilangiocarpic development) :傘蓋邊緣延伸向下卷把子實層包圍。
v. 柄生被果型的發育模式
(stipitoangiocarpic development) :菌柄衍生組包圍子實層但從包圍整個原基。
vi. 球莖被果型的發育模式
(bulbangiocarpic development) :由球包圍整個原基和衍生組包圍子實層。
vii. 被果型的發育模式 (endocarpic development) :子實層被擔子包圍或保護在內,圖示三種成熟腹菌擔子果的直切面。
另外, Reijnders 也提出以下詞匯: 不規則子實層 (rupthymenial)
的分化是遠離中央菌柄的, 光滑子實層 (levhymenial)的菌褶分化是向中央菌柄的, 而裂子實層(schizohymenial)
的則由基原組織分化分裂而成菌褶空間位置 (Volvariella)。除此以外, 菌褶與菌柄尖相連也有8種款式以上。
Reijnders (1948, 1963, 1979) 提出應注意子實體生長的:
1. 內、外菌幕和菌蓋皮層、皮層基質和生長, 還有與子實層體關係,
2. 菌柄、菌蓋和子實層體的發育次序;
3. 子實層體的發育模式。
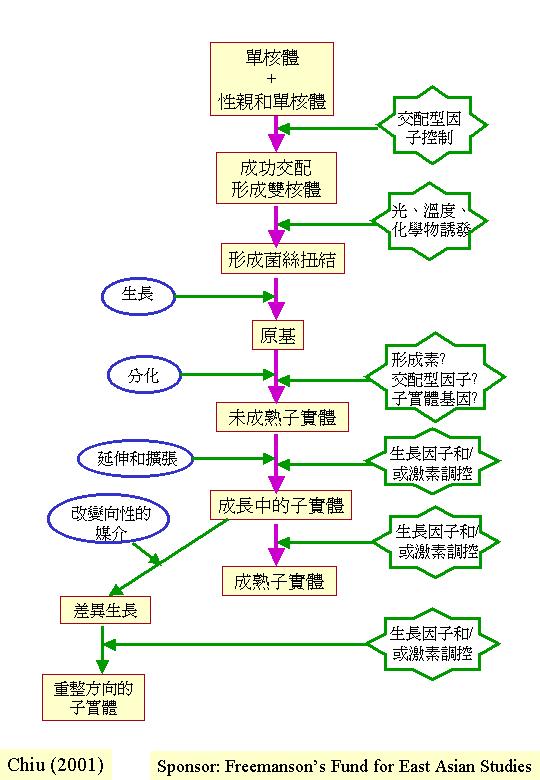
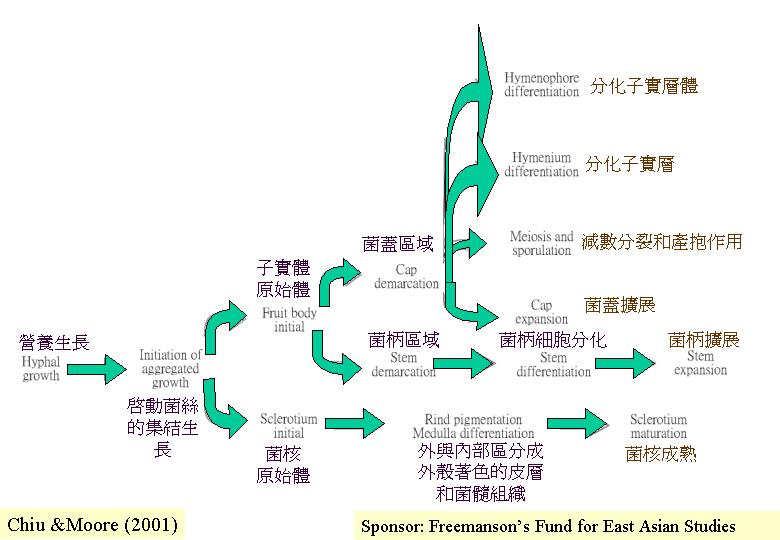
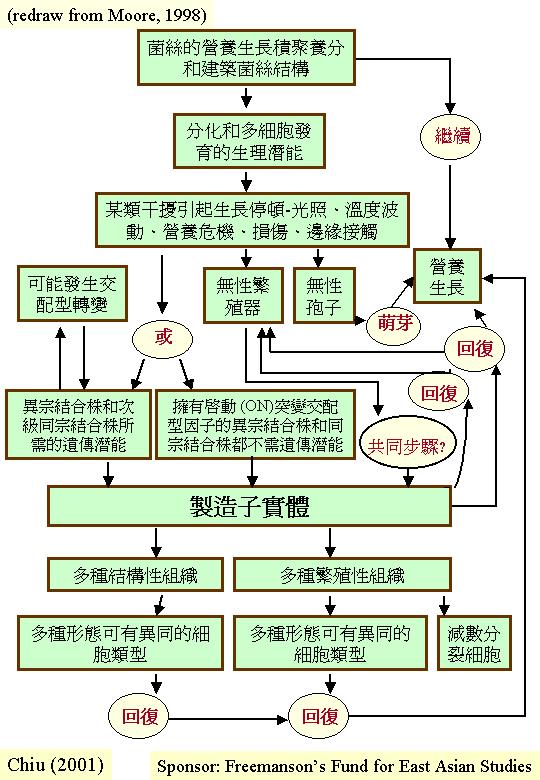
2. 形態的發育
子實體或有關組織 (如:越冬的菌核)由原基發育成長,當中的改變包括細胞類型、分配模式、形狀發展、組織界定和子實體建築的物理和機械原理。 發育其實是動態的過程, 圖示只能表達二度空間,但生長發育是四度空間的:立體的形態隨時間而改變 (Coprinus
fruiting morphogenesis)。
真菌子實體或有關組織的發育不是靠即時的週遭環境獲得資源;菌絲在營養生長時,已將有餘營養轉化囤積留作後用。在適當環境下和擁有合適的遺傳因子, 真菌若擁有充裕的儲存物,便去進行無性生殖或支援產生有性子實體。例:紅曲菌(Monascus
purpureus) 在不同的培養底物下,很快便把外源葡萄糖耗盡,然後在培養瓶內可找到有性和無性的繁殖結構 (Monascus
reproduction modes )。
多細胞的性器官形態發育常是複雜的,有許多平行發展的子程序
(subroutine),各子程序是獨立但又互相影響。子程序的獨立性,可反映在人工培養環境下而出現的部分組織,而非整體器官 (Butler & Wood, 1988)
,又或子實體缺陷型的出現 (閱Chapter 6)。
2a. 發育專一性 (commitment) 、再生
(regeneration) 、重新結實
(renewed fruiting)
從田間抽離的子實體組織可提純菌株,,差不多每部分都有重生
(regeneration) 能力,這是司空見慣的, 但為甚麼這些外殖體不再生子實體,而是倒回到營養菌絲的階段? 若一組織是由天然環境抽離仍肯定地繼續分化, 這就稱為分化的決定
(determination), 但若這切除的組織需要特別的環境因素才可繼續分化, 這就稱為特化(specificaton) 。
Chiu & Moore (1988a) 將不同成熟程度的灰蓋鬼傘菌褶或菌柄 (包括黑暗中長出的菌柄,
Buller, 1931) 切出,然後接種到不同的培養基上 (包括水瓊脂, water agar) ,再觀察外殖體的變化,和重新結實的情況,將結果量化。以Bastouill-Descollonges
& Manachere (1984) 的詞匯形容這重新結實的現象, 發現可有三類模式:
i.
新一代的子實體在外殖體上出現 (名直接結實) ,
ii.
新一代的子實體只在新長出的營養菌絲上出現 (名間接結實),
iii.
是i 和ii 的混合體。
外移殖體會在4天內直接結實, 這比營養菌絲需10-14天結實還要快
(Chiu & Moore,1988a),顯示結實的潛能, 但不代表發育的專一性。結果顯示:只有雙核期的才集中進行直接結實, 減數分裂的菌褶則最小直接結實, Descollonges & Manachere
(1984) 認為「再生現象表明組織有記憶, 而菌褶結實是一自主行為」。
鳳尾菇 (Pleurotus
pulmonarius) 表達極強的重新結實能力;已收成的菇若儲存過長、或敗壞的擔子果上容易重新結實,這些可能是子實體專一的殘留代謝物作祟。
2b. 流動媒體、建築結構與形態發育
真菌的多細胞結構發展需要菌絲合作形成和分化的。生殖器官充滿黏液(圖 6.5;
Van der Valk & Marchant, 1978; McLaughlin, 1982; Williams et al.,
1985; Umar & Van Griensven, 1997a)。體外間質的重要成分是β-1→
6,β-1→ 3 連結葡聚糖 (Dubourdieu & Ribereau-Gayon,1981; Buchala
& Leisola, 1987; Chaubal et al., 1991; Nicole et al., 1993,1995)
、蛋白質合成物和多糖 (Palmer et
al., 1983)。真菌的體外間質可能負責:(i) 識別底物;(ii) 黏附底物;(iii)
修正體外的離子環境;(iv) 儲存、集中和保留體外物質;(v) 抵抗脫水和不利的周圍變化;(vi) 結綁菌絲;和 (vii) 作為或其他效應物 (如:激素) 的運輸媒體。
許多擔子菌散播擔孢子的方法是利用表面脹力引發的彈射作用
(surface tension catapult), 故名彈射孢子
(ballistospores) 或孢子鎗
(spore gun) (Webster et al.,
1995) 。擔子菌中的傘菌其菌蓋形狀會影響空氣流動,因而影響孢子的散播 (Deering
et al., 2001)。
對於擔孢子的釋放,有不同的學說,有說如水玉霉 (Pilobolus) 的黑色孢子囊,靠營養運輸而出現的水脹力 (turgor pressure) ,將整個孢子囊從梗上被強力射出,像擲球般。但 Buller (1901, 1922, 1924) 早期已見到擔子上的孢子梗上有水珠,稱Buller’s droplet,後來的化學分析發現其成分有己糖 (hexose) 和甘露醇
(mannitol) 等吸水
(hydrophilic) 物質,再加上活動過程的拍攝,推論表面脹力才是彈射擔孢子的能源
(Webster et al., 1995)。
子實層體
(hymenophore) 雖有不同模式,但表面通常有水份或流動黏膜包圍,這膜的形成可能是擔子分泌的己糖或甘露醇,引起空氣中水份的吸引和在擔子器上凝固。另外,活躍的呼吸作用和運輸都令到子實體的水分提升,使子實層在穩定而潮濕環境形成、發育和成熟。同樣地,氣流在鈴狀菌蓋(bell-shaped
pileus) 的下方相對菌蓋的週邊是比較靜止的
(Deering et al., 2001) ,這樣,彈射出來的孢子,不易掉在子實層體上,而是離開子實層體
(hymenophore) ,再隨風飄揚。要確保減少擔孢子的浪費,當然還有其它措施 (basidiome)。
Butler & Wood (1988) 研究腐木性的貝狀木層孔菌 (Phellinus contigus) ,發現形成 (多孔菌的) 隔膜 (dissepiment) 的兩個過程是同時進行的, 這兩個過程是: 側生與平生去形成孔管,和趨地性 (gravitropic) 生長去延伸隔膜。 只有培養基是向下放時, 孔才正常出現, 若培養基是向上放時, 孔便不規則生長, 顯示隔膜延伸是正趨地性的。Butler
(1992a)找出隔膜菌絲分化的指標為: 氣生骨架菌絲的趨向生長和分化。Gorovoj et al. (1987) 看到擔子菌的三類模式:
一:除部分例子如班褶菇 (Panaeolus)
外,大部分擁有肉質和生長快的傘菌和多孔菌,其子實層體長時間生產擔孢子,菌褶和孔管的直切面都是狹窄和楔形的,這形狀好讓擔孢子自由放出, 因此, 子實層體表現正趨地性, 有時或會影響菌柄的生長。
二: 鬼傘及其近親等傘菌的菌褶是平行的, 擔孢子同時產生, 而菌褶自溶時, 擔孢子形成和被釋放, Buller (1922)指出鬼傘菌褶是非趨地性的, 若此是事實,菌柄才是負責位置定向的。而且,菌柄的延伸就讓菌蓋或子實層體遠離靜止地面,釋放的擔孢子就有更大機會隨風飄揚。
三: 第三類在那些堅硬的托架菌/栓菌 (bracket fungi)可見到,只有新長出的組織而非整個子實體表現趨地性, 子實體原基是一團未分化組織, 若在直立樹幹出現, 則形成支架, 若在樹枝底部長出, 則邊緊貼樹幹形成圓形,活像一倒轉的菇蕈, 若樹幹受到不規則的干擾, 新的孔維持趨地性的表現, 子實體形卻有極大的變化
(Buller, 1922, 110頁),由此可見, 向性行為可引伸為一形態效應
(Ingold, 1953) 。
2c.
形態發育常受外界誘發
激素和生長因子可影響和協調菌絲尖聚集,在子實體的發育應有許多作用 (Novak
Frazer, 1996), 但是這方面的化學資料實在太少(Uno
& Ishikawa,1982; Wessels,1993; Novak-Frazer, 1996)。化學趨向物導向菌絲生長,這樣可引起菌絲互纏 (Gooday & Adams, 1993;
Gooday, 1994; Mullins, 1994) ,這些化學趨向物有的被描述為性激素
(Bu’Lock, 1976)。 水生的壺菌異水霉 (Allomyces)
產生單鞭毛的雌和雄動配子,但雌配子不甚游動,雌配子和配子囊產生誘雄激素 (雌誘素,sirenin),它或源於焦磷酸法呢酯
(farnesyl phosphate),是雙環的倍半二烯
(sesquiterpenediol) 令雄配子表現強烈的趨向行為 (Nutting et al., 1968; Pommerville, 1981; Pommerville et
al., 1988)。若沒有誘雄激素, 雄配子便隨意打弧、停頓和再不定的打弧游動; 約 10-10 M的誘雄激素已能導向雄配子的游動;若雄配子遠離激素源,它會縮短打弧間的活動量,又若雄配子游向激素源,停頓的數目會減少,與此同時,雄配子會令誘雄激素失活,有關機制是否透過降解或不可逆向地與抑制物結合就不甚清楚
(Machlis, 1973) ,但結果就是雌雄兩配子能接觸。
此外,近以真菌的水霉類綿霉 (Achlya)
的雌性激素名雄器形成激素(antheridiol), 是一類固醇物質
(steroid), 生物測試方法發現其活性濃度為
10-11 M (Raper, 1952; Barksdale, 1969)。雄器形成激素和藏卵器形成激素
(oogoniol) 是一分枝合成途徑的不同產物;雄菌絲受雄器形成激素影響, 擴大導致藏卵器形成激素的合成途徑, 增加呼吸, 誘發葡聚糖的崩解而且觸發纖維素的全程合成。這些代謝變化導致雄器的形成, 包括在管細胞分化點集結小囊 (Mullins & Ellis, 1974;
Horgen, 1981; McMorris, 1978a,1978b ; Timberlake & Orr, 1984)。雌菌絲受藏卵器形成激素影響是有可能產生相似的反應。 雖然其基因機制仍未清晰, 但可參照動物受固醇的影響, 固醇受體位於細胞核, 而激素-受體複合物直接結綁DNA去調控基因表達 (Horgen, 1977)。 然而, 線霉的雄器形成激素有點不一樣
(Horton & Horgen, 1989):激素受體出現在細胞質
(Riehl et al., 1984) ,但其化學構造與其他的類固醇激素受體相似
(Riehl & Toft,1984;
Mullins, 1994)。
當毛霉 (Mucor)
遇到親和的交配型菌絲時﹐孢囊梗的生長受壓制,而性分化則發生(包括製造性菌絲 (接合子幹)的構造), 菌絲向彼此生長、接合、融合而形成配子囊。接合子梗的建築由三孢子酸 (trisporic
acid) 決定;如將三孢子酸添加至純種、未交配的培養中, 菌絲生產接合子梗去取代孢囊梗 (Gooday, 1973b, 1974b; Bu’Lock,
1976; Gooday, 1994)。
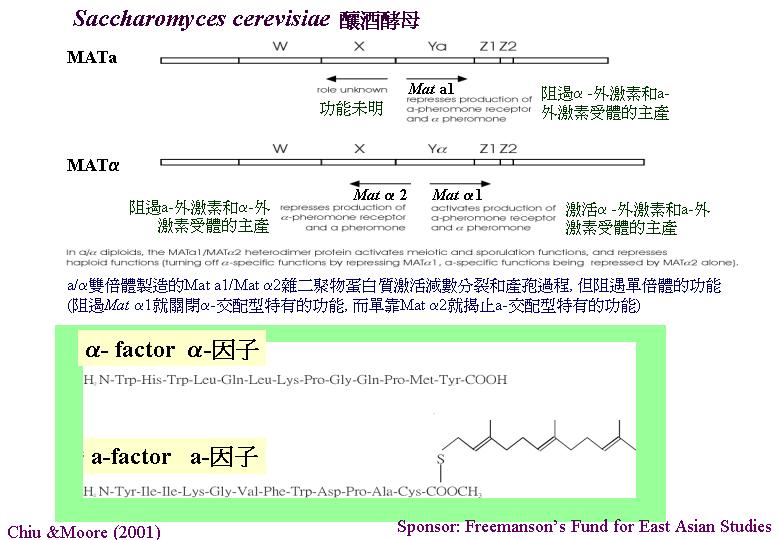
釀酒酵母 (Saccharomyces cerevisiae) 的無性生殖靠萌芽(budding)。性親和的單倍體融合成dumbell-shaped
zygote (合子) ,在營養豐富下,雙倍體進行萌芽成純體系細胞堆 (clone) 。受到饑荒時,雙倍體進行減數分裂,成一四孢子的子囊。子囊孢子的萌動靠萌芽,單倍體在實驗室下進行有絲分裂成單倍純體,性親和的單倍體在自然環境下立即交配,因此,單倍體階段非常濃縮(Chiu & Moore, 1999)。釀酒酵菌交配型基因就是負責製造縮氨酸 (peptide) 激素的(稱為α-和β- 外激素(pheromones); sex factors)
和外激素特定的接受器/受體 (receptor)。酵母的激素與動物界的是異曲同工: 組織交配程序, 所以被命名為外激素,交配程序指: 當外激素接合在親和配子表面的外激素接受器時, 藉著GTP蛋白質作用改變新陳代謝和達致:
(i)
引領接受細胞產生凝集素
(agglutinin) 令兩親和配子黏著;
(ii)
令細胞停留在細胞周期的 G1
階段;
(iii)
改變細胞壁結構和其結果細胞形狀。兩外激素引起它們的目標細胞延長進入接觸範圍, 但是相同的親合細胞或雙倍體則沒有反應 (Chiu
& Moore, 1999)。
玉蜀黍黑粉菌在單細胞單倍體類的像酵母擔孢子期時, 若受到親和的外激素刺激會形成接合管 (Chiu & Moore, 1999)。玉蜀黍黑粉菌的外激素是短的脂: 有由11-15
種氨基酸組成的C-末端, 最後的是一半胱氨酸殘基連接法呢酯基團(15-C
異戊二烯部分),後者令外激素極度疏水。
有些真菌表現歸巢現象 (homing reaction);管狀細胞尖向同種的發芽擔孢子生長(Kemp, 1977; Moore, 1984b) ,雖然有關機制未明。近期對裂褶菌和灰蓋鬼傘的交配型因子B的序列分析,發現類似釀酒酵母的外激素和外激素受體序列 (Kűes & Casselton, 1992)。 既然交配型因子B是控制細胞核的遷徙和鎖狀細胞與母管狀細胞融合(mating
type control), 因此, 交配型因子B的產物(為轉錄因子)估計是管理和導向鎖狀細胞的。導向生長和扭結不一定限於同源,Kemp (1977) 更描述二個不同的鬼傘種 (Coprinus miser和 C. pellucidus)可共同組成一子實體, 若從子實體的各部分(包括子實層、菌柄)抽離組織放在培養基上, 兩種菌絲都再生出來。
許多不同化學品和子實體提取物都能誘發、提高或影響真菌子實體的生產和形態發育, 如: 腦脂類
(cerebrosides) 誘發裂褶菌 (Schizophyllum
commune)結實、雙孢蘑菇 (Agaricus
bisporus)子實體提取物、構巢曲霉 (Aspergillus
midulans)性缺陷種的酚內酯 (phenolic
lactones)和埋核盤菌 (Pyrenopeziza
brassica) 提取物都影響子囊果的發育 (Nelson,
1971; Wolf & Mirocha,1973; Rusmin & Leonard, 1978; Kawai & Ikeda, 1982;
Ilott et al., 1986; Campe & El-Zyat, 1989; Siddiq et al., 1989)
,還有,亞油酸 (linoleic
acid) 作用於長喙殼 (Ceratocystis)
、粗糙孢菌 (Neurospora
crassa) 和叢赤殼 (Nectria
haematococa) ,而真菌產孢吸光色素 (mycosporines) (Dyer et al., 1992) 作用於燒土火絲菌 (Pyronema omphaloides) 、羊肚菌 (Morchella esculenta) 和叢赤殼 (Nectria galligena) 等,亞油酸等是否成形素 (morphogen) 或營養物
(nutrient) ,則文獻未有交待。大部份這些物質都能刺激生長, 但在鬼傘(C. congregatus) 原基期的提取物則是抑制生長的(Robert & Bret, 1987; Robert,
1990)。但它們絕不是激素或生長因子, 因為:
i. 它們都是基本的代謝物, 這不符合激素的專一性;
ii. 其有效濃度遠超訊號分子的特性;
iii. 生物測試 (bioassay) 這些物質只表現有或無的反應, 卻不能像典型成形素般,在濃度的梯度下誘發不同的分化模式,形成一形態 (發育)場(morphogenetic
field)。 Novak Frazer (1996) 指出生長因子必定是內在或受降解而變成不穩定的,因此,可在器官內形成一濃度梯度和分化場,而其作用時間應是短暫的。
2d. 形態模式/ 模型結構、形態場與電腦模擬
縱然廣泛的因素促進子實體的生產, 從紜紜的觀察中可得出三個概論, 試述如下:
首先, 真菌有效及迅速地將環境養料內化, 操縱內在營養的管理、資源分配和調動, 因此,營養菌絲可產生像子實體的多細胞結構。
其次,無論促進子實體因素是物理的或化學的,它們都干擾細胞的正常新陳代謝。干擾本身才是促進子實體成因。 結果, 子實體的發展看起來有許多平行的途徑, 而每一途徑的影響因子又是有關係和可互換的 (如: 某營養條件可用光照取代;底物
NH4+ 濃度可代替光刺激結實) ,應環境調控而縮減增熟某一發育途徑下,這便表現出各式各樣的子實體形態 (fruit body
polymorphism) (Watling, 1971; Chiu et al., 1989, 1999)。
第三, 就算最簡單的生育發展途徑都可細分幾個步驟 (至少包括:形成、發育和成熟), 而且每一步驟都需要訊號(干擾)指引, 因此, 連續的訊號 (連續的變化干擾) 驅使發育程序滾動; 沒有干擾, 序列便停滯下來。 更多複雜構造 (如子實體) 都表明形態發育是複雜和多向的, 並擁有許多平行途徑的, 而菌絲的生理正影響繼續某一途徑或抉擇透過分枝點轉進另一平行途徑(fungal morphogenesis)。
引致模式 (pattern) 出現的成形素如: 著名的視黃酸
(retinoic acid)和植物生長素 (auxin), 視黃酸是維生素A的衍化物, 它在細胞間存在並形成一梯度濃度作為細胞與細胞通訊的橋樑,並引領雞胚胎的足芽分化中層細胞, 產生足指(Brickell and Tickle, 1989; Tickle, 1991), 這就是因應位置的成形素(morphogen)
濃度而作出的反應,因而表達發育向性,視黃酸與細胞核的視黃酸受體結合, 調控許多同源基因 (homeobox genes) (Tickle, 1991),其基因產物就規劃足指的位置。植物生長素同樣透過一濃度梯度指引胚胎的細胞形態和導管細胞的分化
(Sachs, 1991) ,外加的植物生長素透過連接到細胞核的受體活化植物組織, 轉錄特定的 mRNAs去進行細胞延長和分裂
(Key, 1989; Guilfoyle et al., 1993; Löbler and Klämbt, 1985; Guilfoyle et
al., 1993; Ulmasov et al., 1995) , 由此可見, 植物生長激素引起器官專一的、組織專一的和發育專一的表達模式
(Guilfoyle et al., 1993)。
現時只可估計真菌同樣有發育成形素(Moore, 1998) 。如:灰蓋鬼傘 (Coprinus cinereus) 的菌褶組織中心 (gill organizer) 有可能由位置訊息決定, 若兩個中心太接近, 其形態場就抑制第三個出現, 但當菌蓋長大時, 即兩個中心距離加大, 第三個中心便出現, 最後菌褶便形成輻射狀對稱地包圍菌柄的模式 (Rosin and Moore, 1985a;
Moore, 1987)。
Moore (1995, 1996) 提出一假設: 擴散 (diffusion) 的激活劑 (活化訊息物) 帶領菌褶組織中心前進, 而菌褶組織中心也生產一抑制劑阻止新的菌褶組織中心在其擴散範圍內出現, 當菌蓋膨大時, 兩個菌褶組織中心的距離拉遠, 抑制劑作用減弱至消漓, 新的菌褶組織中心便會形成, 因此, 在激活劑和抑制劑的擴散和可作用濃度下,形成一控制菌褶位置、數量和輻射狀排列的菌褶場。
此外,灰蓋鬼傘子實層上囊狀細胞 (cystidia) 的非隨意分佈是另一例 (Horner & Moore, 1987), 光學顯微鏡下, 囊狀細胞是明顯和肥大的細胞 (cystidial
morphogenetic field), 因此, Horner & Moore (1987) 將灰蓋鬼傘菌褶的切片進行數據分析,發現: 同子實層的囊狀細胞配對(cis)
(兩者從相同的子實層出現) 和相向子實層的囊狀細胞配對(trans) (從相向的子實層出現) 不是完全隨意分佈的, 同子實層的囊狀細胞配對比較少出現,暗示在同一子實層有抑制現象影響囊狀細胞的分佈;這說明囊狀細胞有一形態場, 而這形態場的半徑有 30μm, 而且只在同一子實層作用。
當然這囊狀細胞形態場可能是激活劑和抑制劑的相互作用而形成的,
Meinhardt and Gierer (1974) 和
Meinhardt (1984) 就認為激活劑催化自我的合成, 而抑制劑就是壓抑激活劑的合成, 兩者都在生產源頭擴散開去, 不過抑制劑跑得較快, 控制範圍廣, 這些假設,透過包含擴散速度系數、降解速度等的涵數而作的電算機模擬, 就可產生一模擬的形態場 (Meinhardt, 1984), 這模擬可應用在植物葉上的氣孔、胚胎上的纖毛、昆蟲毛髮和刺毛的分佈, 葉紋和葉排列的生長發育上 (Meinhardt, 1984) 。這類電腦模擬形式的好處時它可因應需要而作擴充,最近,
Stočkus & Moore(1996) 採取完全不一樣的策略, 卻能描繪模擬菌柄的向地性彎曲現象 (Moore et al., 2000), 雖然, 那些模擬並不完美, 因為這些數學模擬原是用於植物的, 因而需要附加的其他數項去考慮真菌特質, 另外, 真菌模擬系統仍只是二度空間的, 與植物三度空間的模擬仍有距離 (Korn, 1993)。
2e.
擔子菌菌絲分析
所謂菌絲分析是指將各種細胞分門別纇、統計和應用到分類去。Corner
(1932a, b)用手切片加機器切片去對黃柄小孔菌 (Microporus
xanthopus)子實體作菌絲分析, 他發現四類菌絲:
·
無分枝、厚壁、無鎖狀細胞的骨架菌絲
(sketelal hyphae) ;
·
分枝、薄細胞、側絲有鎖狀連合胞 (clamp connection) 的生殖菌絲
(generative hyphae) ;
·
分枝、厚壁、無鎖狀連合的但紡織其他菌絲的聯絡菌絲
[Pouzar (1966) 使用串連 (ligative) 菌絲這名詞] ;
·
直排、分枝、厚壁、無鎖狀連合的中間菌絲
(mediate hyphae)。
Corner (1932b) 更認為生殖菌絲為基礎, 骨架菌絲在生長區演變而聯絡菌絲在生長區後方發展出來, 所謂中間菌絲是指由生殖菌絲過渡至骨架菌絲的中間類型。三類基本傘肉菌絲系統為:
·
一菌絲型
(monomitic) : 只有一類生殖菌絲。如:珊瑚菌 (Clavaria) (Corner , 1950);
·
二菌絲型
(dimitic) : 生殖菌絲和骨架菌絲或聯絡菌絲。如:許多溝褶菌 (Trogia
spp.) (Corner, 1966);
·
三菌絲型
(trimitic) : 生殖菌絲、聯絡菌絲和骨架菌絲三類菌絲型。如:溝褶菌 (Trogia
stereoides) (Corner, 1966) 。
另一方面,Singer (1986) 和 Kűhner
(1980) 應用菌絲分析在傘菌上,Reijnders
(1977, 1993) 的觀察奠定菇蕈的菌絲分析,Pegler (1995) 總結在腹菌的擔子菌 (馬勃、土星和鬼筆) 見到的各種菌絲。
在電腦模擬下,透過延伸、膨大、分枝和細胞壁的特性等可引起菌絲的不同形態。讀者對菌絲分析要小心,用詞代表研究者所見的形態,菌絲系統並不絕對界限分明, 其實, 菌絲系統和其他形態特徵一樣是人工的分類方法。
2f.
灰蓋鬼傘 (Coprinus
cinereus) 菌柄的形態發育
Reijnders
(1963) 研究了許多擔子果原基,敘述許多生長過程的形態改變, 開始時擔子果原基各部協調生長, 後來局部組織的急促成長, 便出現菌柄延伸和菌蓋擴張 (Coprinus
fruiting morphogenesis)。灰蓋鬼傘(Coprinus cinereus) 擔子的減數分裂卻方便地成為一客觀準確的發育成熟的指標(hymenium
development),還有,這一切都可從一小塊菌褶組織(1
mm2) 去觀察擔子的分化;在釋放孢子前5小時, 菌柄有急速伸延, 在孢子出現時, 菌蓋擴張, 在孢子成熟時, 菌蓋在1小時完成自溶, 因此,在擔子果的各部分出現協調生長 (Coprinus
fruiting morphogenesis),特別是細胞膨大
(cell expansion)。形態發育的研究必須了解和留意生長方向、幾何性和結構性的特點,如鬼傘由1mm
生長至34mm 時, 菌柄周界增加9倍,但菌蓋增長15倍, 結果是體積擴大3000倍。
灰蓋鬼傘的菌柄廷伸可分兩階段, 第一階段的速度較慢, 約10μm
min-1, 第二階段可達110
μm min-1, 即在12小時內有80
mm的菌柄延伸(Moore &
Ewaze, 1976), 多人發現菌柄廷伸主要在菌柄的上半部發生,
Eilers (1974)發現當菌柄伸延時, 細胞長度有68倍的增加和細胞數量有2倍的增加, 同時,
DNA 量增加和細胞變成多核,所以, 菌柄生長的主要機制是細胞延長(Gooday, 1975b; Kamada &
Takemaru, 1977a)。
Hammad
et al. (1993a) 發現減數分裂前的3mm和8mm 高的菌柄細胞直徑沒有加大 , 因此, 此段的生長是靠細胞數量的增加, 其後, 由8mm 延伸至25mm子實體進行減數分裂時, 細胞的橫切面積有加大至6258
μm2, 特別是在菌柄的中、底部, 頂部和極底部的細胞則維持在3000 μm2的水平, 最多細胞延長區域在中上部,當子實體由3mm
加高至83 mm時, 其長/寬比例由2增至35, 因此,這表明灰蓋鬼傘利用細胞延長和菌絲膨大去增大子實體。
灰蓋鬼傘 (Coprinus cinereus) 菌柄的形態發育。27
公釐高的時候,菌柄為實心的,中央的 (區域 4) 和外圍的 (區域 1) 細胞較小,而最大的細胞膨大程度在區域3
(近中央區) 發生;從 27-mm長到70 mm的菌柄時,區域3的細胞直徑有3.6倍的增加。區域2的則有1.6倍的增加。當然,其他位置的細胞嘗試重組去配合區域3的迅速加大,但空間(lumen) 卻在菌柄(中央區)出現(Moore,
1996) (Coprinus
fruiting morphogenesis)。這中空的菌柄減輕體重,但樹立擴張的菌蓋。
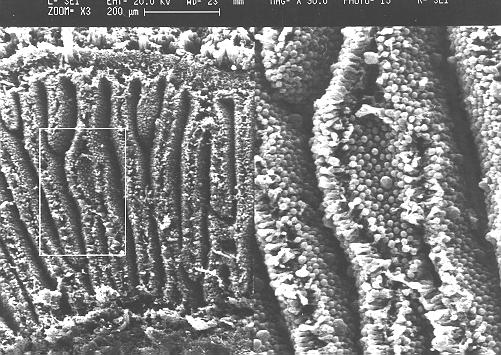
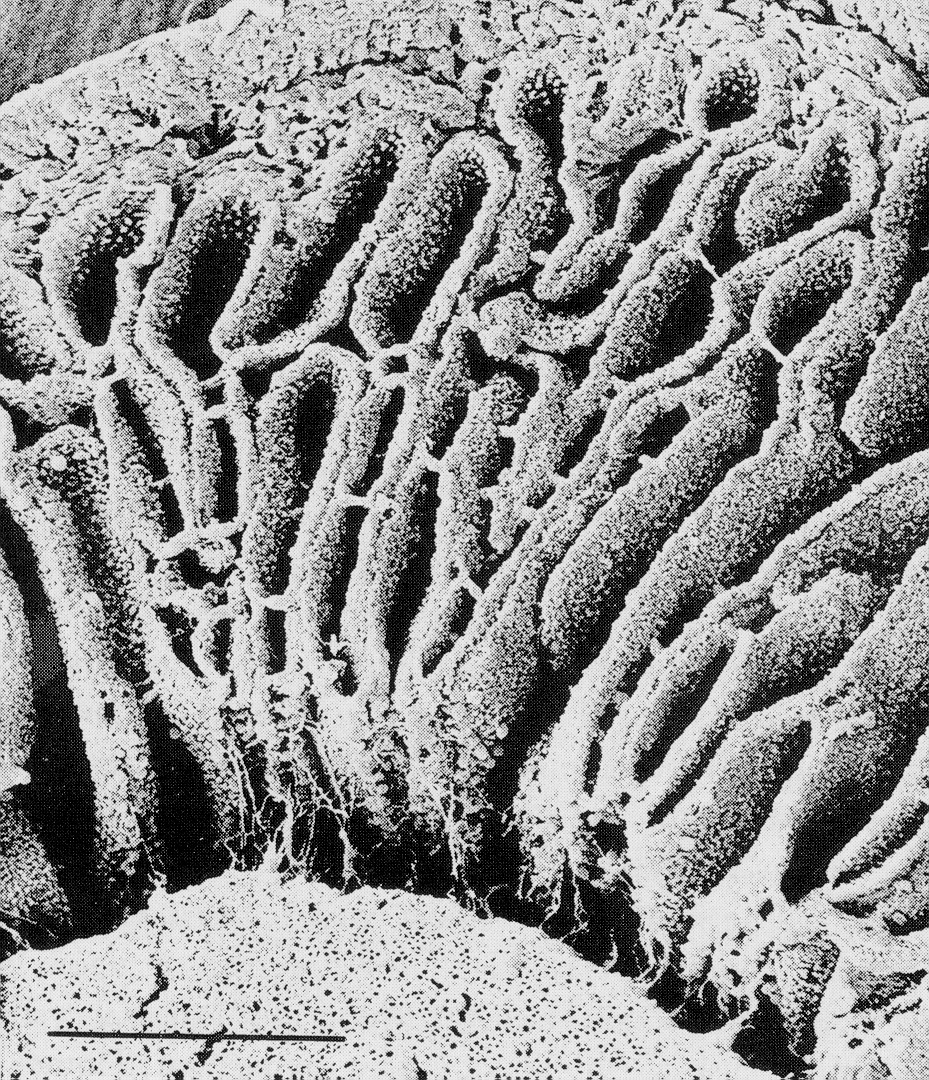
灰蓋鬼傘的菌柄由肥大和延長細胞組成, Lu (1974)描述菌柄中央由雙核菌絲而週遭是多細胞核菌絲組成, Gooday
(1975)用N-乙氨基葡萄糖發現狹窄的菌絲只有頂端有標記,而肥大的菌絲則均稱地各處有標記。在數毫米的菌柄橫切片中大部份是肥大菌絲, 分散的狹窄菌絲只在高倍數顯微鏡下出現, 在直切片和掃描電子顯微鏡照片上都可清晰見到狹窄菌絲,
Hammad et al. (1993b) 將由未成熟到成熟的菌柄作5 μm
橫切片,從外到內作量化菌絲分析 (gravitropism)
, 直徑0-10
和 10-20 μm2
的狹窄和肥大菌絲是最多出現的類別菌絲,將3794細胞分成5μm2為距離的類別作分佈得一線性圖表 , 發現: 狹窄菌絲佔有率為23-54%, 但以面積計算,則只有1-4%, 它們都是有分枝的而且側連到其他細胞去成一網絡, 肥大而多核的菌絲則沒有分枝而又不是一網絡, 在橫切面上肥大菌絲表現平均分佈。當菌柄延伸時, 狹窄菌絲的比例往下調, 約25% 會變成肥大菌絲, 令子實體加大。
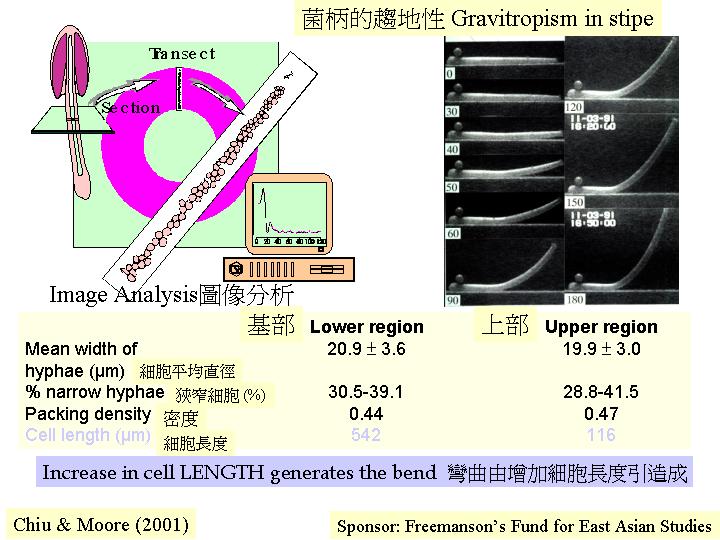
灰蓋鬼傘 (Coprinus cinereus) 菌柄頂的20 至30%
位置表現最大的趨地性彎曲, 再將這部分菌柄分內和外部, Greening et al. (1997) 在光學顯微鏡下觀察,和進行影像分析 (image analysis) (Hammad et
al., 1993a) ,發現:只有細胞長度而非直徑出現分別 (gravitropism), 形成4至5.1與1的差距;若將鬼傘菌柄橫置時,其內圍和外圍長度為1與3.2之比,趨地性彎曲是由外圍細胞的延長而引起;內圍的細胞較小而細胞質包圍隔膜, 外圍細胞長而空泡 (vacuole) 充斥。Kamada
et al. (1991) 發現細胞壁的幾丁質是左旋的,細胞延長可能是加長這幾丁質旋環的距離, 而不加闊細胞。
總論鬼傘菌柄生長包括:部分的狹窄細胞分化成肥大細胞, 肥大細胞延長和加大。這些都表現子實體出現特化區域,而區域內的細胞有不同的分化,最明顯的是:細胞膨大。雖然,不同的功能組織未必有形態分別,但是,真菌多細胞組織中組織膨大是一主要形態發育特徵 (inflation
in morphogenesis )。
2g. 擔子菌菌蓋的形態發育
細胞膨大在許多形態發育的場合出現; 特別是當子實體成熟時 (Reijnders & Moore, 1985), 細胞分化是受即時環境影響的, 正如在粗柄白鬼傘(Leucocoprinus cepaestipes), 表面的菌絲分化成表皮細胞
(pileodermium), 連接的菌絲則分化為外菌幕
(Reijnders, 1948), 又如鬼傘(Coprinus poliomallus)菌褶髓至基原組織餘存的內菌幕(lipsanenchyma) 中的狹窄菌絲就增大變成等圓細胞 (Reijnders, 1979)。
細胞膨大是分化的一種,在子實體原基的生殖菌絲就沒找到膨大的細胞。而膨大細胞的出現就代表原基、菌核和菌根的生長
(Reijnders, 1963) 。連續的相片拍攝和追蹤達13小時,鬼傘菌柄從25 長至93
mm,鬼傘子實體用6小時成長, 菌褶由直向的向外彎而擴展成橫向的
(Buller, 1924; 1931) , 形成開傘的樣子 (Coprinus
fruiting morphogenesis) (Moore et al., 1979) , 雖然Buller
(1931) 提議側絲細胞膨大主要是:分隔擔子,但後來的菌絲分析
(Moore et al., 1979) 發現側絲細胞的出現和膨大導致菌蓋開張, 因為在最後的自溶期間, 擔子雖被分解但側絲細胞仍然存在, 菌蓋的外形保持不變 (Coprinus
fruiting morphogenesis) , 而早期的菌蓋擴張還包括加厚菌肉, 但後期菌肉慢慢變薄,傘肉如一層不能擴張但富彈性的「皮層」(菌蓋皮, pileipellis), 而緊貼下面的菌褶則靠側絲細胞的膨大而引起菌蓋的張開,菌蓋加長而與中央菌柄分開, 於是菌蓋便開展和向上彎 (Coprinus
fruiting morphogenesis) , 因此, 側絲細胞的膨脹是菌蓋開展的主要的機制 (Moore
et al., 1979)。
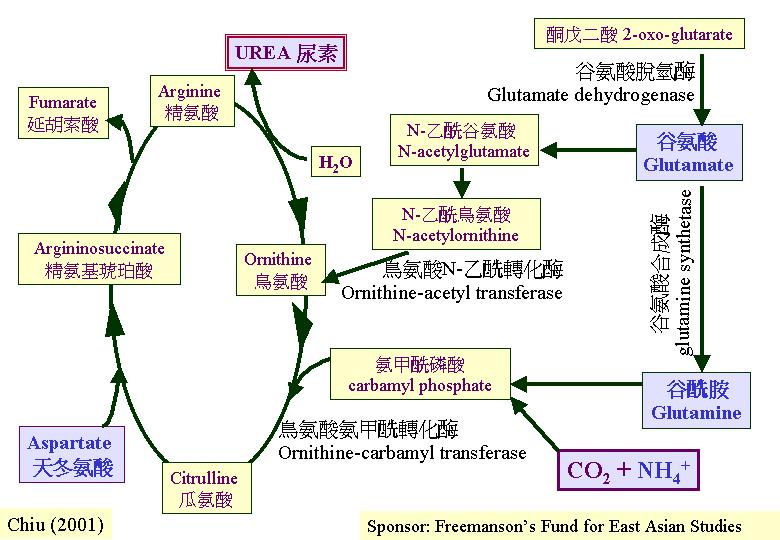
整個子實體就是一機械性結構由壓力、脹力引動形狀的變化。既然膨大的細胞都充滿空泡, 水的引進是極其重要的;大部分傘菌的子實體含水量達70 – 90%之高, 鬼傘利用積聚的尿素(urea)來引起子實體的生長發育(閱Chapter 4) , 同時, 這個尿素是合成餘氮的過程 (urea cycle) 。許多真菌都利用尿素作滲壓劑, 鬼傘菌柄就利用糖複合物, 雙孢蘑菇和香菇就利用達50%體重 (乾重) 的甘露醇
(mannitol),引發水的脹力去支撐子實體。 值得留意: 雙孢蘑菇的菌蓋擴張靠細胞膨大, 香菇就靠增加細胞數量, 顯示同一滲壓劑但不同的擔子果組織擴展的策略。
托架菌、菌 (Bracket
fungi) 子實體許多是長壽的, 因而可釋放更多擔孢子,如:一年生但維持多月的多孔菌(Polyporus)
和多年生的層孔菌(Fomes)。 若仔細研究多孔菌就發現其子實體中有多分枝而纏繞的菌絲去綁結鄰近菌絲,這些菌絲延伸、加厚壁或變成中空的管導去建造一子實體, 擁菌褶的香菇屬 (Lentinus 和Lentinula) 的多孔菌都有類似情況, Ryvardens (1992)就詳述多孔菌菌絲系統的分類。Fayod
(1889)研究菌褶髓的菌絲系統,發現有四類: 左右對稱 (bilateral)、 聚合(convergent)、有規則
(regular)和不規則的 (irregular), 這個特徵可用來作分類。
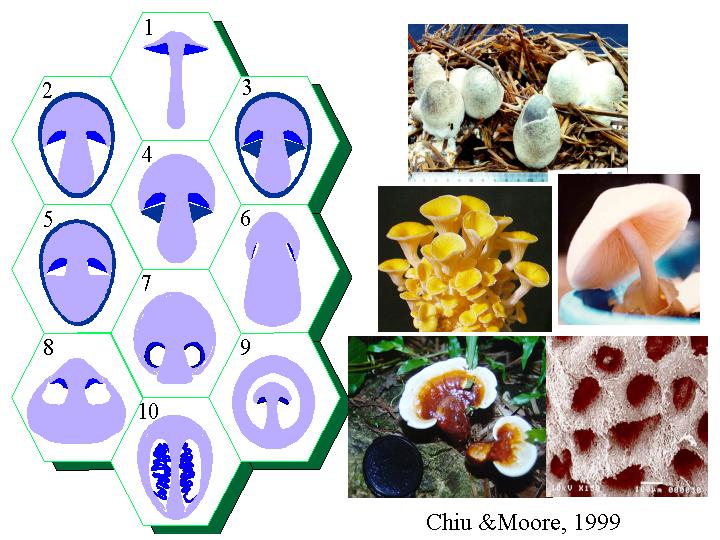
2g -i 子實層體:「胚胎(首)期菌褶」是旋卷的
灰蓋鬼傘的擔子果原基只是成熟的十分一即 800 μm 高時, 已能清晰分辨菌幕
(veil)、菌蓋(pileus, cap)
和菌柄(stipe, stem)
(Moore et al., 1979) (Coprinus
fruiting morphogenesis) 。Reijnders
(1963, 1979) 發現某些真菌沒有完整的柵欄狀細胞層
(palisade layer) 去組成子實層原層, 也沒有菌褶空間 (annular/ gill cavity) 。許多擔子菌的子實層體 (hymenophore) 是以「孤島」(islands)
分散地在菌蓋髓上初現,逐漸地這些「孤島」擴大和連結,形成一種獨特的款式(basidiome
polymorphism) (Chiu & Moore, 1999)。這現象在多孔菌如靈芝和傘菌都得以証明 (Chiu, unpublished result)。
Corner (1932b) 研究黃柄雲芝 (Polystictus
xanthopus) 時, 就提出孔場
(pore field) 的概念, 即菌蓋下部近邊緣處有一菌環 (annulus)
,這裡的菌絲生長、分化形成隴起物和分枝, 再與隔鄰的相連, 形成孔原基
(pore bases) , 菌蓋隔膜的生長就分隔出孔管。Butler
& Wood (1988) 研究腐木性的貝狀木層孔菌 (Phellinus
contigus) 如何在培養瓶上生產平伏的擔子果, 其孔原基在群落邊的後34mm位置出現, 密度與天然的沒分別, Butler (1988) 描述兩個步驟: 島誘發和氣生小簇生長 (小簇
fasicle 指一小朿菌絲) , 島是指菌絲組織互纏成一緊密的子實體原基,這些原基在群落邊緣位置下層菌絲分散地形成, 因此, 發育模式是由位置決定而非菌絲譜系
(Reijnders & Moore, 1985) , 新孔是由這些島原基側生出來的, 其後,氣生小簇生長長出局部寬鬆而生長快速的菌絲, 和側部慢生長或停止生長的菌絲, 雖然隔膜菌絲源自基質, 但其後的局部生長是氣生的, 中間必然由易揮發的新陳代謝物傳送訊息,
Butler (1988) 採用手術實驗去研究有生命的材料, 發現次生孔就在厚的隔膜中長出和將初生孔分裂出來, 慢慢地所有孔的高度都差不多。 這些都反映一孔形態場的情況, 與鬼傘的情況一樣。
初生傘菌菌褶是旋卷如康乃馨;Chiu & Moore (1990a, b) 是第一個在灰蓋鬼傘和銀絲草菇發現初生菌褶是旋卷的。當子實體成長後, 則形成一有規則放射對稱的菌褶子實體, 過程是透過壓力的形成、伸延和消離, 灰蓋鬼傘的子實層上的囊狀細胞就負責這個溝通的功能,銀絲草菇就靠菌褶髓的膨大而去消離這壓力。
2g-ii. 菇蕈造菌褶
菌褶或同等組織是在子實體下方懸掛的,這是否表示菌褶的生長點在開放的一邊? Reijnders和許多真菌分類學家都認為菌褶的生長點是在開放的一邊。但是, 我們以灰蓋鬼傘和銀絲草菇作研究, 發現菌褶的生長發育方向與傳統的看法不一樣。
在傘菌擔子果成長過程中會有更多菌褶出現, 銀絲草菇有兩種產菌褶的方法 (Chiu & Moore, 1990a): 在菌褶的自由邊分叉成二, 然後延伸成兩個新菌褶(making gills) ;另外, 在菌褶根部連接傘肉位置分枝而成第二及第三代的菌褶, 向空間位置伸延開去形成一複雜旋卷的形態。銀絲草菇就靠菌褶髓的菌絲膨大而撐直菌褶的 (Volvariella) , 這就成為有效的輕體重但聳立結構的建築方法。
鬼傘在原基子實體時, 已可清晰看到菌褶, 橫切片下, 菌蓋與中央菌柄形成同心環,菌褶有兩類, 初等菌褶其褶髓是與菌柄相連的, 待成長後菌褶脫離菌柄, 邊緣細胞分化成囊狀細胞 (margin cystidia; Rosin & Moore, 1985a; Moore, 1987; Chiu &
Moore, 1990b); 次等及其他菌褶的子實層是完整複蓋褶邊的。當子實體成長時, 菌柄週界加大, 但菌褶厚度沒增加, 反而菌褶數量增加了。灰蓋鬼傘除擁有銀絲草菇以上兩種的增加菌褶方法 , 還有一獨特的方式;就是在現存的菌褶從褶髓接連菌柄處一分為二, 形成一Y形構造 (Y-shaped gill) ,而且Y的開口往菌肉髓處加大。若從囊狀細胞在子實層隨生長發育的分佈變化看, 也可得到相同結論 (Chiu & Moore, 1990b); 囊狀細胞由平均分佈演成只集中在次生菌褶的邊緣位置。Y形構造的菌褶在雙孢蘑菇 (Agaricus bisporus) 的成長中擔子果也有發現 (Moore, 1998) 。
既然,子實層上有囊狀細胞-接囊狀細胞的組合 (Horner & Moore, 1987) 將隔鄰的菌褶結連起來,初生菌褶又是與菌柄相連的,既然成長中的菌蓋未開, 新增和舊的菌褶局束在一起成複雜旋卷的形態, 因此, 當菌蓋展開時, 左右的壓力就靠囊狀細胞-接囊狀細胞的組合傳遞和平均減弱開去, 結局是形成一有規則放射對稱的子實體, 基因突變種Revoluta就不形成囊狀細胞-接囊狀細胞的組合 (FREE cystidium), 所以成長後子實體仍是旋卷的 (revoluta) (Chiu
& Moore, 1990b)。銀絲草菇(Volvariella
bombycina) 的菌蓋髓由緊密而膨脹的菌絲組成,而子實層是細胞緊密排列和互扣而成。菌褶結構支撐是褶髓的受壓和子實層的張力,當細胞膨大引起的拉力成為機械動力,菌褶便形成平行而直的建築。
掃墨曾應用來研究須霉(Phycomyces)、Lycopodium 和 巨大曲霉 (Aspergillus giganteus) (Castle,
1942; Trinci & Banburt, 1967),我們應用掃墨在銀絲草菇發育異種株的外露擔子果上,就發現:菌褶由0.5cm 長到2.5 cm, 深度由1.5 增至5mm時, 黑點仍保留在菌褶邊位位置, 因此生長點不在開放邊的位置, 同樣地, 菌蓋邊的黑點也沒有移位, 而是黑點的密度降低了, 顯示在整區都有生長發生,但邊沿位置是早有定案的。菌蓋的發育擴大時, 菌褶底部連接菌蓋處才是生長點, 菌褶由菌蓋基原組織分化出來,菌褶開放的一邊是很早確定下來的。
將鬼傘和包腳菇比較, 無論菌褶以何種方式形成, 菌褶的生長點是在底部而非邊緣位置
(Chiu & Moore, 1993; Moore, 1987)。同樣地, 若用活細胞染料(vital stain; 不抑制生長的色素)加在會長菌褶的多孔菌鳳尾菇 (Pleurotus pulmonarius) 上,顏色標記聚集在菌褶邊, 顯示菌褶生長點在菌褶底部, 但就菌蓋而言, 多孔菌香菇 (Lentinula
edodes) 和鳳尾菇其菌蓋邊延續生長, 這與同屬多孔菌的貝狀木層孔菌 (Phellinus conchatus) 吻合 (Butler & Wood, 1988), 即褶狀多孔菌採用多孔菌菌蓋的生長方法, 但子實層體(子實體)既是褶狀, 則捨棄多孔菌的方法, 轉用傘菌模式。
對於複制菌褶,我們提出由「菌褶組織中心」(gill
organiser) 引動的觀念,它引起分化, 形成新的菌褶, 和帶領向菌肉方向前進, 在菌褶空間的兩邊形成新的子實層, 菌蓋的擴張加大這菌褶空間, 菌褶組織中心向菌肉方向推進,它同時促使子實層增長,形成新的皮層和加大菌褶空間,名為制菌褶空間作用 (空間製造cavitaion; gill
organizer; Moore,1987, 1995, 1996), 菌褶組織中心的不斷活動,而以輻射對稱狀向外推進, 引發菌髓基原組織的分化, 從而有更多菌褶出現,而自身產生的生長因子影響而形生一發育形態場
(morphogenetic field)。。
3.
擔子果的建築策略
現代的分類學家都有一套標準包括檢查微構造、外觀、生長模式,再加上DNA序列的考查 (閱Chapter 1),而發現許多真菌是相關的 (Reijnders,1963; Reijnders & Stalpers, 1992;Sugiyama, 1998), 但少數人留意真菌的彈性生長形態 (phenotypic plasticity)。變異應正名為可塑性, 這可有多層次和多個原因下發生, 在實驗室下某一菌株 (strain)可表達子實體形態的多樣性, 也可以是同種異株的分別,前者屬於後遺傳(epigenetic) 的彈性表現,即正常生長發育受到干擾,但基因型沒變化,這可能是適應環境壓力的表現,如雙孢蘑菇的玫瑰巢病
(rose-comb disease), 特徵是菌蓋頂部出現旋卷形生長的子實層, 這正是養菇場內灑噴礦物油導致的
(Lambert, 1930; Flegg, 1983; Flegg & Wood, 1985), 另外, 蜡蘑(Laccaria)、蜜環菌(Armillaria)
和絲蓋傘(Inocybe)等受過濾性病毒感染或真菌攻擊, 都會出現這情況
(Blattn et al., 1971, 1973) ,
Buller (1922) 就描述皮辣乳菇 (Lactarius
piperatus) 受寄生的泌乳菌 (Hypomyces
lactifluorum) 攻擊會出現缺菌褶的子實體,
Watling (1974) 發現粉褶蕈Entoloma
abortivum 子實體原基受蜜環菌影響會長成馬勃狀。
雖然有報導這類子實體的生長彈性(Buller,
1922, 1924; Keyworth, 1942; Singer, 1975), 較詳盡的報告就寥寥可數: 糞生裸蓋菇Psilocybe
merdaria (Watling, 1971; Reijnders,1977)、 雙孢蘑菇
(Worsdell,1915; Atkins, 1950; Reijnders, 1977; Flegg & Wood,1985), 灰蓋鬼傘
(Chiu & Moore, 1990b)、銀絲草菇
(Chiu et al., 1989) 和靈芝
(Chiu et al., 2000)。雙孢蘑菇的子實體變異有: 不育子實體 (carpophoroids; Singer, 1975), 分枝(forking,
bichotomy, Atkins, 1950; Worsdell, 1915), 繁生 (proliferaion; Worsdell, 1915)、連結 (fasciation; Worsdell, 1915), 過多子實層
(Langeron & Vanbreuseghem, 1965), 這些在銀絲草菇 (Chiu et al., 1989) 和糞生裸蓋菇 (Watling, 1971) 都有發現, 還有, 在銀絲草菇也找到羊肚菌狀的 (morchelloid 在鬼傘也遇到,
Chiu & Moore, 1990b) 和似腹菌的 (gasteroid)
, 這些變異子實體和正常子實體同時出現。
Hibbett et al. (1994) 發現虎皮香菇的傘菌狀子實體和似腹菌子實體的早期發育是一樣, 而腹菌狀子實體是因為發育中的菌傘延伸而成。這一見証正衝擊真菌的分類, 既然傘菌可產生類羊肚菌狀的子實體或類腹菌的子實體, 這可能代表返祖現象
(Worsdell, 1915), 而包腳菇屬 (Volvariella)的銀絲草就有可能與腹菌 Brauniella有親緣關係 (Singer, 1955, 1963, 1975), 雖然許多人用個體發育推斷系統發育, 如MacBride’s 胚胎學專書Textbook
of Embryology (1914), 但另一些動物學家如 Walter Garstang’s 則持相反意見: 個體發育不包含系統發育, 而是創造系統發育
(Garstang, 1922)。
銀絲草菇的異形子實體是突發產生的;銀絲草菇沒受寄生蟲、疾病和有毒反應誘發這些突變而來的子實體。 兩個品種在不同的培養基下產生半被果的、被果的和裸果的子實體, 而裸果的子實體的菌托
(volva)可以是缺乏、加厚和正常薄膜狀的, 單生和聚生的子實體可以是:不育的、腹菌狀的、正常傘菌狀的和過多子實層的, 而子實層體由波狀、旋卷狀的形態至類羊肚菌的和正常傘菌狀的, 某些子實體更見到子實層的變異, 擔孢子被濃濃的黏膜包圍、同一擔子上的孢子不同步地成熟、擔孢子數量變異和擔孢子由卵形變成心形等。 既然兩個種都表現這情況, 正好反映一個體發育的程序,實由各自獨立調控但互相協調的步伐連串而成
(Chiu et al., 1989), 重要的是: 所有子實層都保持產孢能力, 因此, 發育的彈性就是在惡劣環境下,提高產孢子和有利釋放孢子的能力。
真菌的分化可被看為各個階段交叉途徑的連串決策 (如subroutines), 而許多例子都顯示: 真菌保持在某一分化階段內, 但並不硬性固定在某一階段, 而是容許不同程度的表達, 所以所謂決策, 其實代表未知數, 有某一程度的或然率,
如灰蓋鬼傘的菌褶上的囊狀細胞通常是雙核的, 但間中可遇到一巨大單核 (Chiu & Moore, 1993) 或帶有擔孢子梗的形式。 至於囊狀細胞長出菌絲,
Watling 早在1971年研究 糞生裸蓋菇(Psilocybe
merdaria) 一異種子實體時便有發現,若利用抑制劑也可引菌褶外殖體上的擔子出現同樣現象
(Chiu & Moore, 1988a,b; 1990a), 所以,細胞的分化不是牢固的,需要不斷的內在環境發出訊號去維持, 若抽離子實體環境細胞便立刻出現營養生長。Chiu &
Moore (1993)將真菌細胞分化與電腦術語快思邏輯fuzzy
logic (模糊邏輯)聯繫 (Zadeh, 1996), 表示整個系統容許不準確性, 「和/或」的語匯變成「可能」, 簡單來說, 因為輸入資料可有誤差, 結局的線圖擴大了其複蓋範圍, 即在真菌生長發育時容許不準確性。所以, 形態發育不是一連串的決策步驟而引起固定的組合模式, 而是各個小步驟決策的平衡結果, 這樣的快思邏輯可解釋真菌發育的彈性和形態不同的漸變闊度。
鳴謝:
承蒙遠東及香港區共濟會之菲裘新研究東亞學科基金的贊助,使這部介紹真菌生理、生化、遺傳、細胞學、發育和形態及生態的資料可以中文出版,著者深致謝意! 特別鳴謝W. Bro. Peter J. Nunn 先生的支持。
The authors are deeply grateful
to The Freemasons’ Fund for East Asian Studies by the District Grand Lodge of
Hong Kong and the Far East, without which this publication would not have been
possible. We are especially grateful to Mr. W. Bro. Peter J. Nunn, District
Grand Secretary of the Fund for the kind support.
參考書目Reference
Buller,
A. H.R. (1909) Researches on Fungi. Vol. 1. Longmans, Green and Co.:
London. 274 pp.
Buller,
A.H. R. (1922) Researches on Fungi. Vol. 2. Longmans, Green and Co.:
London. 492 pp.
Buller,
A.H. R. (1924) Researches on Fungi. Vol. 3. Longmans, Green and Co.:
London.
Deering,
R., F. Dong, D. Rambo & N. P. Money (2001) Airflow patterns around
mushrooms and their relationship to spore disposal. Mycologia 93(4),
732-736.
Lorenz,
M.C. & Heitman, J. (1998) The MEP2 ammonium permease regulates pseudohyphal
differentiation in Saccharomyces cerevisiae. EMBO Journal 17,
1236-1247.
Moore,
D. (1998) Fungal Morphogenesis. Cambridge University Press.
Webster,
J., Davey, R.A., Smirnoff, N., Fricke, W., Hinde, P., Tomos, D. &Turner,
J.C.R. (1995) Mannitol and hexoses are components of Buller’s drop. Mycological
Research 99, 833-838.
If
you use diagrams or material from this website, we ask you to cite the home
page and authors or the appropriate source publication in your work.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}