
14.14 Direct penetration of the host cell wall
Not all appressoria form in response to stomatal ridges or over stomata; some form over the surfaces of intact cells, and this type of appressorium directly penetrates the host cell wall. Direct penetration is not exclusively mechanical, but extremely high pressures are developed (Money, 1999). The appressorium develops as a swelling at the tip of the germ tube hypha, and is bonded to the host with a strong adhesive. Melanin is deposited within the appressorium wall as it matures, and as this happens the appressorium accumulates metabolites, such as glycerol, which increase the osmotic potential of the cytoplasm. This draws water into the appressorium, generating extremely high turgor pressure.
Melanin deposition in the cell wall of the appressorium is essential for maintaining this high turgor pressure; melanin becomes highly cross-linked into the wall to sustain the physical pressure resulting from the high osmotic pressure.
Next, a thin hypha, called a penetration peg, extends from the base of the appressorium and perforates the host surface. The full force of the turgor pressure built up in the appressorium is focussed in the minute contact area at the tip of this penetration peg and, in addition, hydrolytic enzymes specific to components of the plant cuticle are secreted from the penetration peg as it passes through the cuticle, being induced by cuticle materials. With penetration achieved, a haustorium is often formed from which hyphae branch and spread though the leaf tissue.
Specific examples of this type of host penetration occur in the powdery mildews caused by Blumeria (old name = Erysiphe) species; and the rice blast pathogen Magnaporthe oryzae. Interestingly, the grey mould pathogen Botrytis cinerea (in the Ascomycota), which is also responsible for the ‘noble rot’ of wine grapes, forms an appressorium that does not generate the same physical pressures and lacks the highly melanised wall and the septum that separates the appressorium from the germ tube.
The powdery mildews form an economically important group of plant pathogens that infect a range of plants, including barley, wheat, pea, apple, sugar beet and grapes. The earliest event in powdery mildew infection is the secretion of cutinases and other proteins by the conidia in response to contact with the hydrophobic leaf surface. Within 24 hours from arrival on the plant surface, powdery mildew conidia germinate, develop appressoria, form penetration pegs, penetrate the host outer epidermal cuticle and cell wall, and establish haustorial complexes within epidermal cells. The haustorial complex is the fungal feeding structure that allows the pathogen to draw nutrients from the host cell.
Several transporters, including hexose transporters and amino acid permease, have been identified in the (fungal) plasma membrane of Blumeria graminis (wheat powdery mildew) haustoria, demonstrating that glucose and amino acids are transported from wheat to the disease fungus. The flow of nutrients from the host supports continued hyphal growth over the leaf surface (and many more penetration events). Infected host cells enclose fungal haustoria in an extrahaustorial membrane (EHM), which keeps it separate from the host cytoplasm. The EHM is a unique, specialised membrane of host origin: it might be an invagination of the plasma membrane which is subsequently differentiated, or be synthesised de novo by vesicle trafficking targeted on the invading haustorium. The events involved in a typical powdery mildew infection process are illustrated in Fig. 8 (Koh et al., 2005).
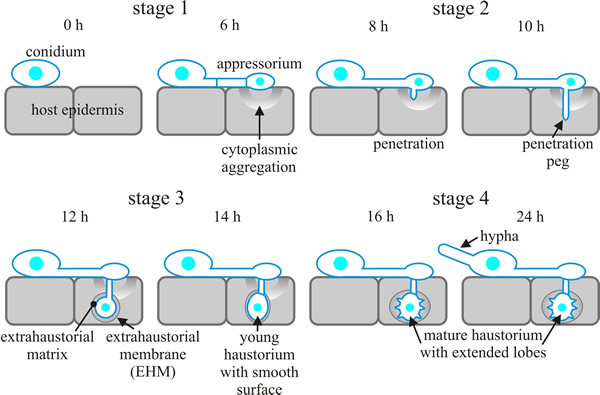 |
| Fig. 8. 'Comic strip' diagrams of the infection sequence of Blumeria (Erysiphe) cichoracearum on a susceptible Arabidopsis host. In stage 1 (0-6 hours after inoculation; all timings are approximate) conidia germinate and develop appressoria, but no penetration into the plant cell occurs. Cytoplasm and organelles of the plant epidermal host cell move towards and accumulate underneath appressoria at this stage. Stage 2 (6-10 h) is the period in which the appressorium developed a long, thin penetration peg, which subsequently penetrates the epidermal cell. Stage 3 (10-14 h) is the stage in which the haustorium first forms at the end of the penetration peg as a swollen, elongated sac with a smooth surface. At this stage, nuclear migration from the hypha on the leaf surface into the haustorium completes, and a septum separates the body from the neck. The haustorial complex, which consists of the fungal haustorium, the gel-like extrahaustorial matrix (which may be of both fungal and host origin) and the host extrahaustorial membrane (EHM) (which is of host origin), develops within 12-18 h. The EHM encases the haustorium, separating it from the host cytoplasm. The last stage of early infection, stage 4 (14-24 h), is characterised by a haustorium with distinct lobes emanating from the body. By stage 4, also, a hypha has emerged from the conidium at the opposite side to the appressorial germ tube. The illustrated sequence of events is representative of other powdery mildews, except that cereal powdery mildews produce a vestigial primary germ tube prior to an appressorial germ tube. Based on illustrations in Koh et al., 2005. |
Approximately 21% of the genes (a total of 2,154) in the genome of Magnaporthe oryzae showed differential expression (changes in gene expression ≥ 2-fold) the majority being up-regulated, during spore germination and appressorium formation (Oh et al., 2008).
During appressorium formation specifically, 357 genes were differentially expressed (240 being up-regulated and 117 down-regulated). There was a significant decrease in expression of genes involved in protein synthesis, while expression of genes associated with protein and amino acid degradation, lipid metabolism, secondary metabolism and cellular transportation exhibited a dramatic increase; there being a four-fold enrichment for genes encoding secreted proteins (Oh et al., 2008)(and see Fig. 13).
Updated July, 2019