
4.3 The fungal lifestyle: colony formation
Following germination, the extension rate of the germ tube increases towards a maximum linear rate, at which point the hypha attains a linear extension rate. The maximum rate of hyphal extension varies greatly between different fungal species and is also dependent on environmental conditions such as temperature, pH and nutrient availability. For example, the extension rate of leading hyphae of Penicillium chrysogenum is about 75 µm h-1 at 25°C, whilst those of Neurospora crassa grown at 37°C can extend at up to about 6,000 µm h-1.
Before the maximum rate of extension is attained, a lateral branch is formed to produce a new growing hypha the extension rate of which also accelerates towards the maximum. As the germling continues to grow, new lateral branches form at an exponential rate and in a pattern that produces the characteristic morphology of the mycelium (Figs 2-5).
 |
| Figs 4.2 to 4.5. These hand-drawn pictures illustrate the well ordered, young mycelium which colonises solid substrates effectively and efficiently. Figs 4.3 to 4.5 show a very young germling below and a slightly more mature colony above. Fig. 4.2 = Mucor (S = position of the germinated spore); Fig. 4.3 = Boletus; Fig. 4.4 = Tricholoma; Fig. 4.5 = Amanita. Fig. 4.2 is redrawn from Trinci, 1974; Figs 4.3 to 4.5 redrawn after Fries, 1943. |
Although individual hyphae in the developing mycelium eventually attain a maximum linear rate, the overall growth of the mycelium is exponential (we will illustrate these features later, and also show how the measurements can be generalised algebraically). During early growth, nutrients surrounding the young mycelium are in excess and the mycelium is unrestricted and undifferentiated. During undifferentiated growth, the mean rate of hyphal extension is dependent on the specific growth rate of the organism (the maximum rate of growth in biomass per unit time) and the manner and degree of branching.
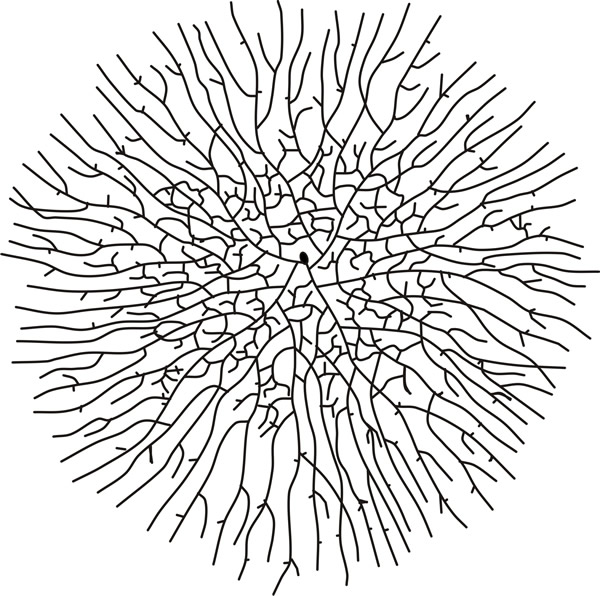 |
Fig. 6. A maturing fungal colony. Notice how the growing hyphae are oriented outward into uncolonised regions whilst the production of branches and hyphal fusions centrally ensures the mycelium becomes a network that efficiently exploits available substrate. This hand drawn sketch of Coprinus sterquilinus comes from one volume of A.H.R. Buller’s epic series Researches on Fungi (Buller, 1909-1934). |
In the older mycelium (Fig. 6) hyphal fusions are evident at the colony centre, and hyphal avoidance reactions at the colony margin. Between them, the sketches in Figs 2-6 portray the main growth processes that influence the distribution of hyphae in a mycelium, which are:
- polarised hyphal growth;
- branching frequency;
- autotropism (the ‘self-avoidance’ reaction that makes vegetative hyphae grow away from the already-existing mycelium).
It is important to recognise that these are dynamic relationships. They change with the age and with the developmental state of the mycelium as its biological functions change, indeed the distribution of biomass in a fungal colony varies with the age of the hyphae. One part of a mycelium may be growing as a rapidly extending, sparsely branched exploratory sector, another part may be a highly branched and interconnected network exploiting a nutrient resource, while a third region reverses the autotropism so that hyphal tips congregate and co-operate in formation of a fruiting structure.
Updated July, 2019