
2.7 Fossil fungi
Most fungal structures are very poor candidates for preservation over long periods of time as fossils. Fungal hyphae are microscopic and have so few unique morphological features that it has been difficult to establish much of a fossil record for fungi and there is always considerable debate over the fungal nature of the materials studied except in a few favoured rock formations (Taylor et al., 2015; Edwards et al., 2018; Krings et al., 2018). Nevertheless, filamentous fossils in vesicles and fractures of a 2.4-billion-year-old basalt from the oldest Proterozoic (Ongeluk) Formations in South Africa, that branch, anastomose and interweave like fungal mycelium have been claimed to satisfy all the criteria for fossilised fungi. Though the authors point out that:
‘…The Ongeluk fossils, however, are two to three times older than current age estimates of the fungal clade. Unless they represent an unknown branch of fungus-like organisms, the fossils imply that the fungal clade is considerably older than previously thought, and that fungal origin and early evolution may lie in the oceanic deep biosphere rather than on land…’ (Bengtson et al., 2017).
Loron et al. (2019) investigated the chemical composition of the walls of isolated multicellular microfossils preserved in shale of the Grassy Bay Formation of Arctic Canada. FTIR spectra were typical of chitin (N-acetyl-D-glucosamine) and/or chitosan (deacetylated-glucosamine). This, together with morphological (resemblance to fungal sporangia) and ultrastructural (layered walls) features indicate that the microfossils (called Ourasphaira giraldae) have a fungal affinity. The Grassy Bay Formation dates to approximately 1 to 0.9-billion years ago:
‘…These microfossils are more than half a billion years older than previously reported unambiguous occurrences of fungi, a date which is consistent with data from molecular clocks for the emergence of this clade. In extending the fossil record of the fungi, this finding also pushes back the minimum date for the appearance of eukaryotic crown group Opisthokonta, which comprises metazoans, fungi and their protist relatives.’ (Loron et al., 2019).
Bonneville et al. (2020) used what they describe as ‘…an array of microscopic (SEM, TEM, and confocal laser scanning fluorescence microscopy) and spectroscopic techniques (Raman, FTIR, and XANES)…’ to demonstrate the presence of vestigial chitin in fossil filaments found in a 810 to 715 million year old shale from the DR Congo. These authors firmly identified the fossil filaments and mycelium-like structures as remnants of fungal networks, representing the oldest, molecularly identified remains of kingdom Fungi, and conclude:
‘…the fossil filaments resemble more Blastocladiomycota hypha. … our study represents the oldest, documented fungi to date and pushes well into the Neoproterozoic the possibility that fungi helped to colonize land surface, almost 300 [million years] before the first evidence of land plants. Overall, our discoveries also lend support to previous assumptions regarding the role of fungi in land colonization and, by extension, on the evolution of Earth’s biogeochemical cycles.’ (Bonneville et al., 2020).
 |
|---|
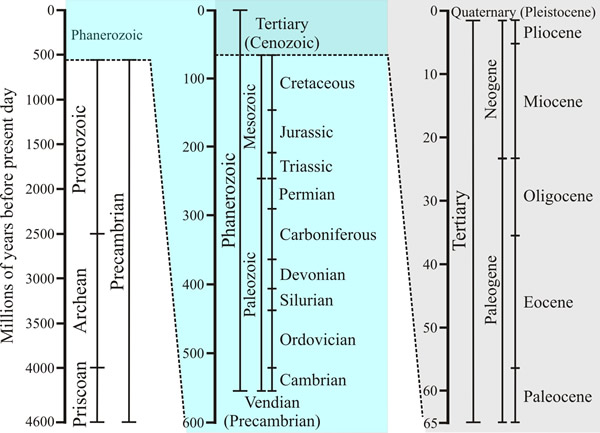
| Fig. 7. The geological time scales. The figure shows how the entire span of time during which the Earth has existed can be classified into eras and epochs. Time axes on the central and right-hand panels are magnified views of the upper section of the previous panel. The last 65 million years is called the Cenozoic era, and is divided into several epochs, which began with the Palaeocene approximately 65 million years ago and runs to the present day. The Palaeocene, which lasted from 54 to 65 million years ago is followed by the Eocene (34-54 million years ago), the Oligocene (24-34 million years ago), the Miocene (5 to 24 million years ago), the Pliocene (1.8 – 5 million years ago) and the Pleistocene or Ice Ages (10,000 years ago to 1.8 million years ago). The period since the last retreat of the glaciers and present glacial warming is called the Holocene, or Recent (up to 10,000 years ago). Source (and see for more details): https://en.wikipedia.org/wiki/Geologic_time_scale (page edit of January 2018). |
Some of the oldest terrestrial 'body' fossils of any sort are large fibrous things called nematophytes. They are part of what is known as ‘phytodebris’ and provide the earliest evidence for land organisms. They have been found from the mid-Ordovician (460 million years ago; see the geological time scale in Fig. 7) to the early Devonian, suggesting that they lasted a period of at least 40 million years. This phytodebris certainly contains fossils of bryophyte-like plants, but it has been suggested that some of the nematophytes (particularly the genus Prototaxites) were terrestrial fungi (Hueber, 2001).
Two remarkable things about these fossils are that they were extremely large (Fig. 8), and so common that they were a major component of these early terrestrial ecosystems, both in terms of abundance and diversity. They included by far the largest organisms present in these ancient environments. Indeed, ‘…specimens of Prototaxites over a metre wide have been reported...’ (Wellman & Gray, 2000).
Two remarkable things about these fossils are that they were extremely large (Fig. 8), and so common that they were a major component of these early terrestrial ecosystems, both in terms of abundance and diversity. They included by far the largest organisms present in their ancient environments. Indeed, ‘…specimens of Prototaxites over a metre across have been reported...’ (Wellman & Gray, 2000). They are described as ‘Devonian fossil logs’ and have been conjectured to be kelp-like aquatic algae, rolled up carpets of liverworts, giant lichens or enormous saprotrophic fungal fruiting bodies.
 |
Fig. 8. A Lower Devonian Prototaxites
compression fossil, at least 2 m tall, in situ in the Bordeaux
Quarry, Quebec. Dr Francis Hueber, who first made the suggestion that
Prototaxites fossils are fungal in origin (Hueber, 2001), is posing
alongside as a scale marker. Photograph from Boyce et al.,
2007.
What you are looking at here is a stream bed that has been upended by
geological movements to be at right angles to its original position. So,
although this is now a vertical rock (quarry) wall, you are effectively
now looking down on the stream bed. |
|---|
Boyce et al. (2007) measured the ratio between different carbon isotopes to determine the nature of the fossils. The argument runs that a photosynthetic primary producer will have a relatively constant ratio of carbon isotopes between individuals because they are all using atmospheric carbon. On the other hand, organisms (like fungi) that are classified as consumers will take on the isotope ratios of whatever they are digesting locally, and therefore individual specimens end up with widely differing isotope ratios. Boyce et al. (2007) found too much isotopic variance between individual Prototaxites fossils for them to be photosynthetic primary producers. Instead, Prototaxites was a consumer, and taken together with direct microscopic observation of their anatomy (Hueber, 2001) this demonstrates that these enormous fossils, the largest land organisms to have lived up to their point in time, were giant fungi. Another interesting factor is that the isotope ratios of Prototaxites fossils can indicate the sorts of primary producers available for its mycelium to digest, and several Prototaxites individuals tested come from environments that just did not include the vascular plants that dominate the modern world. Even though conquest of the land by vascular plants had begun about 40 million years prior to the emergence of Prototaxites. Instead, the Prototaxites environment was still dependent on the more ancient primary producers: cyanobacteria (blue-green algae), eukaryotic algae, lichens and mosses, liverworts, and their relatives (bryophytes).
Retallack & Landing (2014) examined a 386-million-year-old fossil of Prototaxites loganii, a specimen known as the ‘Schunnemunk tree’ (as it comes from the Middle Devonian Bellvale Sandstone on Schunnemunk Mountain, eastern New York). This specimen was 8.83 metres long and had six branches on the upper 1.2 metres of the main axis, each branch being about 1 metre long and 9 cm in diameter. The histology of this specimen is different from that expected of algae, liverworts or vascular plants. The authors claim it best resembles a lichen tissue with coccoid phycobionts, and though Prototaxites has previously been placed within Basidiomycota (as a sort of giant club-fungus), they consider the organisms may belong within Mucoromycotina or Glomeromycotina.
So the current understanding is that the first really large terrestrial organisms were large multicellular fungi that developed to take advantage of 2 billion years worth of accumulated bacterial, protist and bryophyte debris (Fig. 9).
 |
|---|
Fig. 9. Two artistic
impressions of the Lower Devonian landscape of some 400 million years
ago, dominated by specimens of Prototaxites up to 9 metres
tall. Top is a painting by Mary Parrish of the Smithsonian Institution,
Washington, which was prepared for the publication about
Prototaxites fossils by Hueber, 2001. At bottom is a painting by
Geoffrey Kibby that appeared under the title ‘an artist's impression of
the landscape of the Devonian period’ as a rear cover image on the
magazine Field Mycology in April 2008. In the landscape
portrayed in these paintings the fungus Prototaxites dominates
as the largest terrestrial organism to have lived up to this point in
time. Although vascular plants had emerged by this time, these
landscapes were still dependent on the more ancient primary producers:
cyanobacteria (blue-green algae), eukaryotic algae, lichens and mosses,
liverworts, and their bryophytes relatives. What you are seeing here is
the physical expression of the dominance of fungi in the Earth’s
biosphere. This physical dominance of Prototaxites lasted at
least 40 million years (about 20 times longer than the genus Homo
has so far existed on Earth). Images kindly supplied by Tom Jorstad of
the Smithsonian Institution and Geoffrey Kibby, senior editor of
Field Mycology. |
Many of the most convincing fungal fossils are associated with plant fossils, including Glomeromycotan (used to be called glomalean) mycorrhizas, and ascomycete or chytrid parasites. Among the most ancient of these are from the Devonian Rhynie Chert of Aberdeenshire in the north of Scotland (400 million years old) in which mycorrhizal fungi (recognisably from the Glomeromycotina) and several other fungi have been found associated with the preserved tissues of early vascular plants (Taylor et al., 2015 [see their chapter 7, Glomeromycota, pp. 103-128]; Edwards et al., 2018; Krings et al., 2018; Berbee et al., 2020).
Glomeromycotan fossils have also been found in mid-Ordovician rocks of Wisconsin (460 million years old). The fossilised material consisted of entangled, occasionally branching, nonseptate hyphae and globose spores. The age of these fossil Glomeromycotan fungi indicates that such fungi were present before the first vascular plants arose, when the land flora most likely only consisted of bryophytes, lichens and cyanobacteria. Today, the Glomeromycotina form the arbuscular mycorrhizal symbiosis, which is ubiquitous in modern vascular plants and has also been reported in modern hepatics and hornworts. As the ancient fungi were present both prior to the emergence of vascular plants and in the tissues of early vascular plant fossils it seems reasonable to suppose that arbuscular mycorrhizas played an important role in the success of early terrestrial plants (Redecker et al., 2000). ). Certainly, genomic analyses have revealed strong congruence between the evolutionary histories of the oldest lineages of living vascular plants with their fungal communities (Benucci et al., 2020).
Other convincing fossils are more recent. Microfossils of hyphae with clamp connections are known from the Pennsylvanian/Carboniferous (300 million years ago; see Fig. 7). However, the only convincing mushroom fossils found so far are preserved in amber dating from the Cretaceous (about 90 million years ago). These ‘mushrooms in amber’ (Hibbett et al., 1995) are particularly interesting because they bear a strong resemblance to the existing genera Marasmius and Marasmiellus, which are quite common in modern woodlands; yet when they were preserved the dinosaurs still ruled the Earth. In other words: the mushrooms you see when you trek through the forest are almost identical to those seen by dinosaurs in their forests. Although, of course, the forest plants are very different.
Amber dated to the Eocene (54-34 million years ago; see the geological time scale in Fig. 7) has been found that contains the remains of several filamentous mould fungi. Among the finds are sooty moulds in European amber dating back to 22-54 million years ago. Present day sooty moulds are a mixed group of saprotrophs, usually with dark coloured hyphae, which produce colonies superficially on living plants (as harmless epiphytes). Most present day sooty moulds use arthropod excretions for nutrition and live closely associated with aphids, scale insects and other producers of honeydew. All the fossils are composed of darkly coloured hyphae with features identical to the present day genus Metacapnodium, suggesting that Metacapnodium hyphae have remained unchanged for tens of millions of years (Rikkinen et al., 2003). Possibly the most impressive fossil is a piece of amber from the Baltic region that contains an inclusion of a springtail (a collembolan arthropod) which is overgrown by an Aspergillus species (Dörfelt & Schmidt, 2005). The surface of the springtail is densely covered in places by excellently preserved hyphae and conidiophores. Numerous sporulating conidiophores can be seen easily, and conidial heads with radial chains of conidia are clearly visible. As well as superficial hyphae at the cuticle, the springtail is loosely penetrated by branched substrate hyphae, so the authors suggest that the fungus may be parasitic and describe it as a new species, Aspergillus collembolorum (Dörfelt & Schmidt, 2005).
A survey of fleshy fungi from amber deposits around the world found representatives of gilled fungi, puffballs, bird’s nest fungi, gasteroid fungi, xylaroid fungi, hymenomycetes, polypores and included the first fossil morel from amber collected in the Dominican Republic (Poinar, 2016). Another recent review found 137 records of non-lichenised fungi and 182 of lichens; about 10% were macrofungi, the rest microfungi. The finds ranged from the Carboniferous (about 310 million years ago) to the Upper Miocene (about 10 million years ago) (Halbwachs, 2019a & b; 2020). So, overall, fossil evidence shows the fungi to be important members of terrestrial ecosystems up to 500 million years ago, and maybe up to 2 billion years ago.
Molecular phylogenetic evidence suggests that fungi are much older. To begin with, molecular phylogenetic studies tended to use single gene sequences. The most popular in fungi have been the nuclear ribosomal DNA (rDNA) locus, particularly that encoding small-subunit (18S) ribosomal RNA but including the nuclear large ribosomal RNA subunit (nucLSU), mitochondrial rDNAs, and complete or near complete mitochondrial genomes. Single-gene phylogenies may not provide sufficient information nor be truly representative to resolve a fungal phylogeny with sufficient confidence, so broader studies provide better information. Sequences of protein-coding genes can be a problem because it’s difficult to design primers for PCR amplification that can be reliably applied to a wide range of taxa. Also, heterozygous loci in heterokaryons can complicate interpretations.
Updated September, 2020