
2.6 The opisthokonts
The opisthokont clade is a distinct lineage of eukaryote groups which share an ultrastructural identity and have no evident sister group; it comprises all the true fungi including chytrids, microsporidia, collar-flagellate protists (Choanozoa), and Kingdom Animalia (which includes us in the metazoa)(Fig. 2.6).
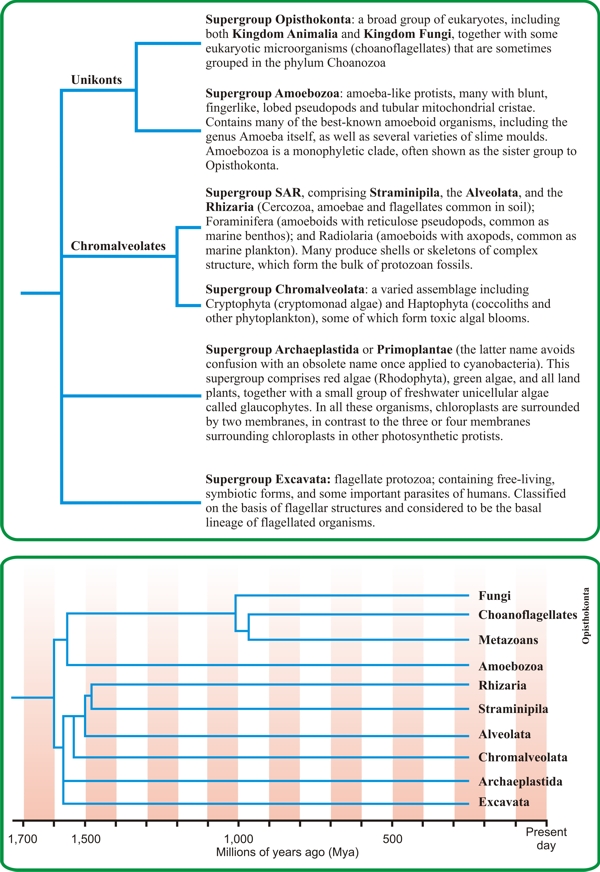 |
| Fig. 2.6. Cladograms (phylogenetic trees) showing the relationships of the five to seven ‘Supergroups’ into which the global tree of eukaryotes is currently subdivided (Adl et al., 2005, 2012; Burki, 2014). In the top panel, the taxonomically formal Kingdoms have been assigned to the deliberately informal supergroups by phylogenomic analysis, a method that uses large alignments of tens to hundreds of genes to reconstruct evolutionary histories. The intention of ‘supergrouping’ is to summarise the relationships arrived at so far while maintaining flexibility as further work on phylogenomics resolves contentious relationships (see: https://www.sciencenews.org/article/tree-life-gets-makeover). The bottom panel shows a simplified version to which a timescale has been added, showing the mean divergence times for the appearance of each lineage. These are shown as minimum (approximate) dates and are based on either body fossil occurrences or molecular fossils (Parfrey et al., 2011; Eme et al., 2014; Loron et al., 2019). |
All molecular and ultrastructural studies have strongly supported the idea that opisthokonts form a monophyletic group (it is informally called Supergroup Opisthokonta; Fig. 2.6, above). The name ‘opisthokont’ comes from the Greek and means ‘posterior flagellum’, so the common characteristic that gives them their name is that flagellate cells, when they occur, are propelled by a single posterior flagellum, and this applies as much to chytrid zoospores as to the sperm of mammals. In contrast, other eukaryote groups that have motile cells propel them with one or more anterior flagella, and that ‘one or more’ phrase results in the name heterokont.
Heterokonts are organisms in which the motile cells have unequal flagella. This is the most nutritionally diverse eukaryote supergroup and includes several ecologically important groupings. The formal name of the assemblage is the Heterokonta and it is placed within the Kingdom Chromista alongside haptophytes and cryptomonads by Cavalier-Smith & Chao (2006). The Chromista seems to represent an independent evolutionary line that diverged from the same common ancestor as plants, fungi, and animals. Heterokonta include:
-
multicellular brown seaweeds, which are the most common type of seaweed on rocky beaches, some forming kelp forests with fronds up to 50 metres long.
-
The usually parasitic oomycetes that include Phytophthora, the pseudofungus that caused the great Irish potato famine of 1845, and Pythium cause of seed rot and damping off in seedlings, which have been confused with fungi because of their filamentous growth form. The inclusion of several groups of ‘pseudofungi’ and ‘water moulds’, especially genera that have been studied long and hard by mycologists over the years (names to look out for are genera like Saprolegnia, Achlya, Albugo, Bremia, Plasmopara), is why the Heterokonta is important to us as mycologists.
-
Numerous protists of major importance, like photosynthetic diatoms, which are a primary component of plankton.
-
Numerous groups of chlorophyll-c-containing algae (chlorophyll-c and a number of other pigments found in the Chromista are not found in any group of true plants).
-
Several non-photosynthetic groups that feed phagotrophically or absorptively.
This is one of the most actively-researched groups of eukaryotes, partly because some biologists doubt that the group is monophyletic, so the components of the group, and its name, are frequently revised (refer to the Tree of Life project website (Keeling et al., 2009). A few people treat the Chromista as identical in composition with the heterokonts, describe them as straminopiles, or seek to change the name of the Kingdom to Straminipila. However, although the name Chromista has nomenclatural precedence (see discussion in Cavalier-Smith & Chao, 2006), we will use the name Straminipila, following Beakes et al. (2014).
Updated March, 2020