
Origins of fungi
The year 1859 was a defining point for Biology. It was the year of publication of The Origin of Species by Charles Darwin, and marked the start of the acceptance of Darwin's proposition that evolution had occurred: that organisms alive today evolved by gradual change from earlier forms. This caused a massive wave of interest in the course of evolution. With animal evolution, study is relatively simple; a great mass of information is already known about morphology of the organisms, and, most important of all, features characteristic to animals, such as bone and teeth, are readily fossilised. However, with fungi, not only is the simple morphology a poor indicator of any past relationships, studies on fossilised fungi have been relatively rare; one important reason being the genuine lack of definitive fossil evidence. Still, this has not prevented many mycologists from phylogenetic speculation relating to the origins and subsequent evolution of fungi. The recent arrival of molecular methods of assessing relationships, with the comparative analyses of molecular sequences (nucleic acid and proteins), has, however, improved the situation massively.
The opisthokonts
The opisthokont clade is a distinct lineage of eukaryote groups which share an ultrastructural identity and have no evident sister group; it comprises all the true fungi including chytrids, microsporidia, collar-flagellate protists (Choanozoa), and Kingdom Animalia (which includes us in the Metazoa)(Fig. 1).
 |
| Fig. 1. A cladogram (phylogenetic tree) showing the relationships of the eukaryotes. Note that the top branch of this tree shows the opisthokont clade as distinct from the other eukaryotic clades. The opisthokont clade subsequently diverges into animals and fungi as sister clades. Adapted from the Tree of Life Project [http://tolweb.org/Eukaryotes]. Figure and caption from Moore et al., 2011 (URL). |
All molecular and ultrastructural studies have strongly supported the idea that opisthokonts form a monophyletic group. The name ‘opisthokont’ comes from the Greek and means posterior flagellum, so the common characteristic that gives them their name is that flagellate cells, when they occur, are propelled by a single posterior flagellum, and this applies as much to chytrid zoospores as to animal sperm. In contrast, other eukaryote groups that have motile cells propel them with one or more anterior flagella (see our Resources Box about the heterokonts).
Resources Box The heterokonts Heterokonts are organisms in which the motile cells have unequal flagella. This group includes several fungus-like organisms that have been studied by mycologists. CLICK HERE to visit a page providing further information. |
Fossil fungi
Most fungal structures are very poor candidates for preservation over long periods of time as fossils. Fungal hyphae have so few unique morphological features that it has been difficult to establish much of a fossil record for fungi (Berbee & Taylor, 2010).
 |
|---|
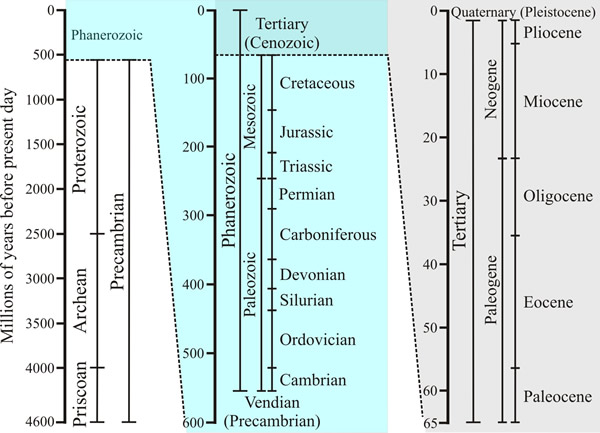
| Fig. 2. The geological timescale. The figure shows how the entire span of time during which the Earth has existed is classified into eras and epochs. Time axes on the central and right-hand panels are magnified views of the upper section of the previous panel. The last 65 million years is called the Cenozoic era, and is divided into a number of epochs, which began with the Paleocene approximately 65 million years ago, and runs to the present day. The Paleocene, which lasted from 65 to 54 million years ago, is followed by the Eocene (54-34 million years ago), the Oligocene (34-24 million years ago), the Miocene (24-5 million years ago), the Pliocene (5-1.8 million years ago) and the Pleistocene or Ice Ages (1.8 million years ago to 10,000 years ago). The period since the last retreat of the glaciers and present glacial warming is called the Holocene, or Recent (10,000 years ago to present day). Source: http://www.geo.ucalgary.ca/~macrae/timescale/timescale.html. Figure and caption from Moore et al., 2011 (URL). |
Some of the oldest terrestrial fossils of any sort are large fibrous things called nematophytes. They are part of what is known as ‘phytodebris’ and provide the earliest evidence for land organisms. They have been found from the mid Ordovician (460 million years ago; see the geological timescale in Fig. 2) to the early Devonian, suggesting that they lasted a period of at least 40 million years. This phytodebris certainly contains fossils of bryophyte-like plants, but it has been suggested that some of the nematophytes (particularly the genus Prototaxites) were terrestrial fungi (Hueber, 2001).
Two remarkable things about these fossils are that they were extremely large (Fig. 3), and so common that they were a major component of these early terrestrial ecosystems, both in terms of abundance and diversity. They included by far the largest organisms present in these ancient environments. Indeed, ‘specimens of Prototaxites over a metre wide have been reported’ (Wellman & Gray, 2000).
 |
| Fig. 3. A Lower Devonian Prototaxites compression fossil, at least 2 m tall, in situ in the Bordeaux Quarry, Quebec. Dr Francis Hueber, who first made the suggestion that Prototaxites fossils are fungal in origin (Hueber, 2001), is posing alongside as a scale marker. Photograph kindly supplied by Dr Carol Hotton of the Smithsonian Institution, appears as Fig 1A in Boyce et al., 2007. Figure and caption from Moore et al., 2011 (URL). |
Boyce et al. (2007) measured the ratio between different carbon isotopes to determine the nature of the fossils. The argument runs that a photosynthetic primary producer will have a relatively constant ratio of carbon isotopes between individuals because they are all using atmospheric carbon. On the other hand organisms (like fungi) that are classified as consumers will take on the isotope ratios of whatever they are digesting locally, and therefore individual specimens end up with widely differing isotope ratios. Boyce et al. (2007) found too much isotopic variance between individual Prototaxites fossils for them to be photosynthetic primary producers. Instead, Prototaxites was a consumer, and taken together with direct microscopic observation of their anatomy (Hueber, 2001) this demonstrates that these enormous fossils, the largest land organisms to have lived up to their point in time, were actually giant fungi.
Another interesting factor is that the isotope ratios of Prototaxites fossils can indicate the sorts of primary producers available for its mycelium to digest, and a number of the Prototaxites individuals tested come from environments that just did not include the vascular plants that dominate the modern world. Even though conquest of the land by vascular plants had begun about 40 million years prior to the emergence of Prototaxites. Instead, the Prototaxites environment was still dependent on the more ancient primary producers: cyanobacteria (blue-green algae), eukaryotic algae, lichens and mosses, liverworts, and their relatives (bryophytes).
So the current understanding is that the first really large terrestrial organisms were large multicellular fungi that developed to take advantage of 2 billion years' worth of accumulated bacterial, protist and bryophyte debris (Fig. 4).
 |
|---|
Fig. 4. Two artistic impressions of the Lower Devonian landscape of some 400 million years ago, dominated by specimens of Prototaxites up to 9 metres tall. Top is a painting by Mary Parrish of the Smithsonian Institution, Washington, which was prepared for the publication about Prototaxites fossils by Hueber (2001). At bottom is a painting by Geoffrey Kibby that appeared under the title ‘an artist's impression of the landscape of the Devonian period’ as a rear cover image on the magazine Field Mycology in April 2008. In the landscape portrayed in these paintings the fungus Prototaxites dominates as the largest terrestrial organism to have lived up to this point in time. Although vascular plants were already present at this time, these landscapes were still dependent on the more ancient primary producers: cyanobacteria (blue-green algae), eukaryotic algae, lichens and mosses, liverworts and their bryophyte relatives. What you are seeing here is the physical expression of the dominance of fungi in the Earth’s biosphere. This physical dominance of Prototaxites lasted at least 40 million years (about 20 times longer than the genus Homo has so far existed on Earth). Images kindly supplied by Tom Jorstad of the Smithsonian Institution and Geoffrey Kibby, senior editor of Field Mycology. The painting by Mary Parrish courtesy of and © Smithsonian Institution. Reproduced with permission from Elsevier. Figure and caption from Moore et al., 2011 (URL). |
Many of the most convincing fungal fossils are associated with plant fossils, including glomeromycotan (used to be called glomalean) mycorrhizas, and ascomycete or chytrid parasites. Among the most ancient of these are from the Devonian Rhynie Chert of Aberdeenshire in the north of Scotland (400 million years old) in which mycorrhizal fungi (recognisably from the Glomeromycota) and several other fungi have been found associated with the preserved tissues of early vascular plants (Taylor et al., 2004, 2006).
Glomeromycotan fossils have also been found in mid-Ordovician rocks of Wisconsin (460 million years old). The fossilised material consisted of entangled, occasionally branching, non-septate hyphae and globose spores. The age of these fossil glomeromycotan fungi indicates that such fungi were present before the first vascular plants arose, when the land flora most likely only consisted of bryophytes, lichens and cyanobacteria. Today, the Glomeromycota form the arbuscular mycorrhizal symbiosis, which is ubiquitous in modern vascular plants and has also been reported in modern hepatics and hornworts. As the ancient fungi were present both prior to the emergence of vascular plants and in the tissues of early vascular plant fossils it seems reasonable to suppose that arbuscular mycorrhizas played an important role in the success of early terrestrial plants (Redecker et al., 2000).
Other convincing fossils are more recent. Microfossils of hyphae with clamp connections are known from the Pennsylvanian/Carboniferous (300 million years ago; see Fig. 1). However, the only convincing mushroom fossils found so far are preserved in amber dating from the Cretaceous (about 90 million years ago). These ‘mushrooms in amber’ (Hibbett et al., 1995) are particularly interesting because they bear a strong resemblance to the existing genera Marasmius and Marasmiellus, which are quite common in modern woodlands; yet when they were preserved the dinosaurs still ruled the Earth. In other words: the mushrooms you see when you trek through the forest are almost identical to those seen by dinosaurs in their forests. Although, of course, the forest plants are very different.
Amber dated to the Eocene (54-34 million years ago; see the geological timescale in Fig. 1) has been found that contains the remains of several filamentous mould fungi. Among the finds are sooty moulds in European amber dating back to 54-22 million years ago. Present-day sooty moulds are a mixed group of saprotrophs, usually with dark-coloured hyphae, which produce colonies superficially on living plants (as harmless epiphytes). Most present-day sooty moulds use arthropod excretions for nutrition and live closely associated with aphids, scale insects and other producers of honeydew. All the fossils are composed of darkly coloured hyphae with features identical to the present-day genus Metacapnodium, suggesting that Metacapnodium hyphae have remained unchanged for tens of millions of years (Rikkinen et al., 2003). Possibly the most impressive fossil is a piece of amber from the Baltic region that contains an inclusion of a springtail (a collembolan arthropod) which is overgrown by an Aspergillus species (Dörfelt & Schmidt, 2005). The surface of the springtail is densely covered in places by excellently preserved hyphae and conidiophores. Numerous sporulating conidiophores can be seen easily, and conidial heads with radial chains of conidia are clearly visible. As well as superficial hyphae on the cuticle, the springtail is loosely penetrated by branched substrate hyphae, so the authors suggest that the fungus may be parasitic and describe it as a new species, Aspergillus collembolorum (Dörfelt & Schmidt, 2005).
Further information about the taxonomy, phylogeny and evolution of fungi can be found in the new textbook 21st Century Guidebook to Fungi by David Moore, Geoffrey D. Robson & Anthony P.J. Trinci. Published 2011 by Cambridge University Press, ISBN: 9780521186957. URL: http://www.cambridge.org/gb/knowledge/isbn/item6026594/?site_locale=en_GB. View Amazon page. |
Updated December 15, 2016